Lipidy
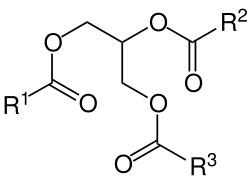
Lipidy (z řeckého lipos – tučný) jsou přírodní látky živočišného i rostlinného původu, mezi které patří především tuky, oleje, vosky, některé vitamíny a hormony. Mohou být v kapalném nebo pevném skupenství.
Chemicky jsou to převážně estery vyšších mastných kyselin a alkoholů. Přesněji řečeno se jedná o deriváty mastných kyselin jednosytného nebo trojsytného alkoholu. Mastné kyseliny jsou vyšší karboxylové kyseliny nasycené nebo nenasycené.

Společnou vlastností lipidů je jejich hydrofobní charakter, který je podmíněný obsahem delšího nepolárního uhlovodíkového řetězce. V biologii a mikrobiologii jsou lipidy makro molekulami, které jsou nerozpustné ve vodě a rozpustné v nepolárních rozpouštědlech. Nepolární rozpouštědla jsou většinou uhlovodíky používané k rozpuštění jiných přirozeně se vyskytujících molekul uhlovodíkových lipidů, které se nerozpouštějí nebo se obtížně rozpouštějí ve vodě, včetně mastných kyselin, vosků, sterolů, vitamínů rozpustných v tucích (jako jsou vitamíny A, D, E a K), monoglyceridy, diglyceridy, triglyceridy a fosfolipidy.
Jedna z funkcí lipidů je ukládání energie, signalizace a působení jako strukturální složky buněčných membrán.[1][2] Lipidy mají také uplatnění v kosmetickém a potravinářském průmyslu i v nanotechnologii.[3]
Vědci někdy definují lipidy jako hydrofobní nebo amfifilní (látky dobře rozpustné v tucích a vodoodpudivé) malé molekuly. Amfifilní povaha některých lipidů jim umožňuje tvořit struktury, jako jsou vezikuly, multilamelární/unilamelární lipozomy nebo membrány ve vodném prostředí. Biologické lipidy pocházejí zcela nebo částečně ze dvou odlišných typů biochemických podjednotek: ketoacylových a izoprenových skupin.[2] Díky tomuto přístupu lze lipidy rozdělit do osmi kategorií: mastné kyseliny, glycerolipidy, glycerofosfolipidy, sfingolipidy, sacharolipidy a polyketidy (odvozené z kondenzace ketoacylových podjednotek); a sterolové lipidy a prenolové lipidy (odvozené z kondenzace isoprenových podjednotek).[2]
Lipidy jsou pro živé organismy nepostradatelné. Měly by tvořit maximálně 25–30 % denní dávky živin pro dospělé. Celkový příjem energie a tuků závisí na pohlaví, věku, hmotnosti a fyzické zátěži jedince. Při denním energetickém příjmu 8500 kilojoulů by množství přijatých tuků odpovídalo 70g.
Historie
V roce 1815 Henri Braconnot klasifikoval lipidy do dvou kategorií a to tuhé tuky či lůj a tekuté oleje. [4] V roce 1823 vyvinul Michel Eugène Chevreul podrobnější klasifikaci, zahrnující oleje, tuky, lůj, vosky, pryskyřice, balzámy a těkavé oleje (nebo esenciální oleje).[5][6][7]
První syntetický triglycerid zaznamenal Théophile-Jules Pelouze v roce 1844, kdy vyrobil tributyrin zpracováním kyseliny máselné glycerinem v přítomnosti koncentrované kyseliny sírové. [8] O několik let později Marcellin Berthelot, jeden z Pelouzeových studentů, syntetizoval tristearin a tripalmitin reakcí analogických mastných kyselin s glycerinem v přítomnosti plynného chlorovodíku při vysoké teplotě.[9] V roce 1827 William Prout uznal tuk spolu s bílkovinami a sacharidy jako důležitou živinu pro lidi a zvířata. [10][11]
Po jedno století považovali chemici za „tuky“ pouze jednoduché lipidy vyrobené z mastných kyselin a glycerolu, ale v průběhu let byly popsány nové formy lipidů. Theodore Gobley (1847) objevil fosfolipidy v mozku savců a slepičích vejcích. Thudichum objevil v lidském mozku některé fosfolipidy, glykolipidy a sfingolipidy.
Pro lipidy měl každý autor trochu jiný název, objevovali se termíny jako např. lipoid, lipin, lipide a lipid. [12] V roce 1912 navrhli Rosenbloom a Gies nahrazení „lipoidu“ „lipinem“.[13] V roce 1920 zavedl Bloor novou klasifikaci pro „lipoidy“: jednoduché lipoidy (tuky a vosky), složené lipoidy (fosfolipoidy a glykolipoidy) a odvozené lipoidy (mastné kyseliny, alkoholy, steroly).[14][15]
Slovo lipid, které etymologicky pochází z řeckého λίπος, lipos 'tuk', zavedl v roce 1923 francouzský farmakolog Gabriel Bertrand.[16]
V roce 1947 definoval T. P. Hilditch „jednoduché lipidy“ jako tuky a vosky (pravé vosky, steroly, alkoholy).
Dělení lipidů
| Dělení lipidů | ||
|---|---|---|
| Jednoduché lipidy | acylglyceroly | Alkohol tvoří glycerol:
|
| vosky | Alkoholem není glycerol, ale například cetylalkohol, myricylalkohol – příkladem může být včelí vosk, nebo ovčí lanolin, často také pokrývají plody rostlin. | |
| Složené lipidy | sfingolipidy | Ceramidy, fosfosfingolipidy a glykosfingolipidy (cerebrosidy a gangliosidy). |
| glykolipidy | Galaktolipidy, sulfolipidy a glykosfingolipidy. | |
| lipoproteiny | Chylomikrony, HDL, VLDL, LDL. | |
| fosfolipidy | Tvoří dvojvrstvu buněčné membrány. | |
| Odvozené lipidy | steroidy, karotenoidy, lipofilní vitaminy a prostaglandiny. | Jsou to látky lipidické povahy, které ale nemohou být svojí strukturou zařazeny mezi lipidy. |
Vznik a složky lipidů
Vznik lipidů
Chemicky jsou lipidy estery vyšších mastných kyselin a alkoholů. Přesněji řečeno se jedná o deriváty mastných kyselin jednosytného nebo trojsytného alkoholu. Chemická reakce vzniku lipidu a vzorec triacylglyceridu:

Složka lipidů – alkoholy
První složkou lipidů jsou alkoholy. V tucích je to trojsytný alkohol glycerol. Ve voscích je to primární nebo sekundární alifatický alkohol.

Složka lipidů – mastné kyseliny
Druhou složkou lipidů jsou mastné kyseliny, vyšší monokarboxylové kyseliny. Mastné kyseliny lze dělit podle různých kritérií – například podle délky řetězce nebo nasycenosti vazeb.
Nasycené mastné kyseliny neobsahují v řetězci žádnou dvojnou vazbu. Tvoří dlouhé přímé řetězce. Zvláště v živočišných tucích je velké množství nasycených mastných kyselin jako energetická rezerva. Příklady nasycených mastných kyselin:
- kyselina kaprylová (CH3(CH2)6COOH)
- kyselina kaprinová (CH3(CH2)8COOH)
- kyselina laurová (CH3(CH2)10COOH)
Nenasycené mastné kyseliny obsahují ve svém řetězci jednu dvojnou vazbu. Zvláště v rostlinách je velké množství nenasycených mastných kyselin. Příklady nenasycených mastných kyselin:
- kyselina palmitoolejová (CH3(CH2)5CH=CH(CH2)7COOH)
- kyselina olejová – cis izomer (CH3(CH2)7CH=CH(CH2)7COOH)
- kyselina eruková (CH3(CH2)7CH=CH(CH2)11COOH)
Chemické složení lipidů
Jednoduché (homolipidy)
- Tuky a oleje – estery vyšších karboxylových kyselin (nejčastěji z kyselin olejové, linolové a palmitové) a trojmocného alkoholu glycerolu,
- Vosky – estery vyšších alifatických kyselin (nejčastěji z kyselin laurové, myristové a palmitové) a primárních nebo sekundárních alifatických alkoholů.
Složené (heterolipidy)
- Fosfolipidy – estery kyseliny fosforečné a různých derivátů glycerolu nebo sfingosinu na jedné své straně, také glycerolu nebo cholinu, ethanolaminu nebo serinu na druhé a zároveň obsahují i dlouhé nepolární mastné kyseliny. Výsledkem je pak, že fosfolipid má polární a nepolární konec,
- Glykolipidy – obdobné fosfolipidům, jen na sfingosin nebo glycerol je glykosidicky navázán oligo- nebo mono-sacharid, který pak v celém lipidu představuje polární konec,
- Lipoproteiny – jsou shluky lipidů s proteiny na svém povrchu. Jsou součástí buněčných membrán, cytoplazmy a krevní plazmy.
- Sfingolipidy – obsahují základní skelet sfingoidních bází – sadu alifatických aminoalkoholů, která zahrnuje sfingosin. Byly objeveny v extraktech mozku a byly nazvány podle mytologické sfingy. Mají význam pro nervovou tkáň.
Biologické funkce

Součást buněčných membrán
Eukaryotické buňky se vyznačují kompartmentalizovanými organelami vázanými na membránu, které provádějí různé biologické funkce. Glycerofosfolipidy jsou hlavní strukturální složkou buněčných membrán, jako je buněčná cytoplazmatická membrána a intracelulární membrány organel; v živočišných buňkách cytoplazmatická membrána fyzicky odděluje intracelulární složky od extracelulárního prostředí.
Glycerofosfolipidy jsou amfipatické molekuly (obsahující hydrofobní i hydrofilní oblasti), které obsahují glycerolové jádro spojené se dvěma "ocásky" odvozenými od mastných kyselin pomocí esterové vazby a do jedné "hlavové" skupiny fosfátovou esterovou vazbou. Zatímco glycerofosfolipidy jsou hlavní složkou buněčných membrán, jiné "neglyceridové" lipidové složky, jako je sfingomyelin a steroly (hlavně cholesterol v membránách živočišných buněk) se v buněčných membránách objevují také.[17]
U rostlin a řas jsou galaktosyldiacylglyceroly[18] a sulfochinosyldiacylglycerol,[19]které postrádají fosfátovou skupinu, důležitými složkami membrán chloroplastů a příbuzných organel a jsou nejhojnější lipidy ve fotosyntetických tkáních, včetně tkání vyšších rostlin, řas a určitých bakterií.
Membrána je formou lamelární fáze lipidové dvojvrstvy. Tvorba lipidových dvojvrstev je energeticky preferovaný proces, když jsou výše popsané glycerofosfolipidy ve vodném prostředí. Toto je známé jako hydrofobní efekt. Ve vodném systému se polární hlavy lipidů zarovnají směrem k polárnímu vodnému prostředí, zatímco hydrofobní ocasy minimalizují svůj kontakt s vodou a mají tendenci se shlukovat a vytvářet vezikuly; v závislosti na koncentraci lipidu může tato biofyzikální interakce vést k tvorbě micel, liposomů nebo lipidových dvojvrstev.
Skladování energie
Triglyceridy, uložené v tukové tkáni, jsou hlavní formou ukládání energie jak u zvířat, tak u rostlin. Jsou hlavním zdrojem energie při aerobním dýchání, protože uvolňují dvakrát více energie kyslíku než uhlohydráty, jako je glykogen, na hmotnost; je to způsobeno relativně nízkým obsahem kyslíku v triglyceridech. [20] Úplná oxidace mastných kyselin uvolňuje asi 38 kJ/g (9 kcal/g), ve srovnání s pouze 17 kJ/g (4 kcal/g) při oxidativním štěpení sacharidů a bílkovin. Adipocyt neboli tuková buňka je navržena pro nepřetržitou syntézu a odbourávání triglyceridů u zvířat, přičemž odbourávání je řízeno hlavně aktivací hormonu citlivého enzymu lipázy.[21] Stěhovaví ptáci, kteří musí létat na dlouhé vzdálenosti, aniž by jedli, používají jako palivo pro své lety triglyceridy.
Další funkce
Objevily se důkazy, že lipidová signalizace je životně důležitou součástí buněčné signalizace.[22][23][24][25] Lipidová signalizace může nastat prostřednictvím aktivace receptorů spojených s G proteinem nebo jaderných receptorů.
Vitamíny „rozpustné v tucích“ (A, D, E a K) – což jsou lipidy na bázi isoprenu – jsou základními živinami uloženými v játrech a tukových tkáních s rozmanitou škálou funkcí. Acyl-karnitiny se podílejí na transportu a metabolismu mastných kyselin do az mitochondrií, kde podléhají beta oxidaci. [26]
Polyprenoly a jejich fosforylované deriváty také hrají důležitou transportní roli, v tomto případě transport oligosacharidů přes membrány.
Polyprenolfosfátové cukry a polyprenoldifosfátové cukry fungují v extracytoplazmatických glykosylačních reakcích, v extracelulární biosyntéze polysacharidů (například polymerace peptidoglykanu v bakteriích) a při N-glykosylaci eukaryotického proteinu.[27][28]
Kardiolipiny jsou podtřídou glycerofosfolipidů obsahující čtyři acylové řetězce a tři glycerolové skupiny, které jsou zvláště hojné ve vnitřní mitochondriální membráně.[29][30] Předpokládá se, že aktivují enzymy zapojené do oxidativní fosforylace. [31]
Lipidy tvoří také základ steroidních hormonů.[32]
Vlastnosti a využití
- Přestože jsou lipidy funkčně i chemicky navzájem velmi různorodé látky, jejich společnou charakteristickou vlastností je hydrofobnost – jsou tedy nerozpustné ve vodě. Naopak se dobře rozpouštějí v mnoha organických rozpouštědlech.
- Jsou energeticky velmi bohaté (1 gram tuku obsahuje 39 kJ = 9,3 kcal), a proto v organizmu slouží jako zdroj a zásoba energie. V lidském těle jsou ukládány ve formě tukových buněk (adipocytů), které slouží jako nejhodnotnější zdroj energie.
- Mnohé druhy lipidů jsou jako lipidová dvouvrstva součástí buněčných membrán, které jsou nepropustné pro vodu. Tvoří i jiné stavební složky,
- Mají ochranné a izolační funkce a slouží i jako rozpouštědlo některých lipofilních látek – především vitamínů. Rozpouštějí vitamíny A, D, E a K, aby je mohl organismus využívat.
- Lipidy mají ochrannou funkci na úrovni celého organizmu – tvoří obal některých orgánů. Jako tělesná izolační vrstva chrání tělo mechanicky a teplotně.
- Tvoří části transportních lipoproteinů a myelinové pochvy neuronů.
Metabolismus lipidů
Hlavními dietními lipidy pro lidi a jiná zvířata jsou živočišné a rostlinné triglyceridy, steroly a membránové fosfolipidy. Proces metabolismu lipidů syntetizuje a odbourává zásoby lipidů a produkuje strukturní a funkční lipidy charakteristické pro jednotlivé tkáně.
Biosyntéza
Když je u živočichů nadbytek sacharidů ve stravě, tak se přebytek sacharidů přemění na triglyceridy. Jedná se o syntézu mastných kyselin z acetyl-CoA a esterifikaci mastných kyselin při výrobě triglyceridů, což je proces zvaný lipogeneze.[33]
Mastné kyseliny vznikají syntézami mastných kyselin, které polymerují a následně redukují jednotky acetyl-CoA. Acylové řetězce v mastných kyselinách se prodlužují cyklem reakcí, které přidávají acetylovou skupinu, redukují ji na alkohol, dehydratují ji na alkenovou skupinu a poté ji opět redukují na alkanovou skupinu. Enzymy biosyntézy mastných kyselin se dělí do dvou skupin, u živočichů a hub se všechny tyto reakce mastných kyselin provádí jediným multifunkčním proteinem, zatímco u rostlinných plastidů a bakterií provádějí jednotlivé enzymy jednotlivé kroky v dráze. Mastné kyseliny mohou být následně přeměněny na triglyceridy, které jsou zabaleny v lipoproteinech a vylučovány z jater.
Syntéza nenasycených mastných kyselin zahrnuje desaturační reakci, při které je do mastného acylového řetězce zavedena dvojná vazba.
Syntéza triglyceridů probíhá v endoplazmatickém retikulu metabolickými cestami, při kterých se acylové skupiny v mastných acyl-CoA přenášejí na hydroxylové skupiny glycerol-3-fosfátu a diacylglycerolu.
Terpeny a isoprenoidy, včetně karotenoidů, jsou vyrobeny sestavením a modifikací isoprenových jednotek darovaných z reaktivních prekurzorů isopentenylpyrofosfátu a dimethylallylpyrofosfátu. Tyto prekurzory mohou být vyrobeny různými způsoby. U zvířat produkuje tyto sloučeniny acetyl-CoA, zatímco u rostlin a bakterií se využívají jako substráty pyruvát a glyceraldehyd-3-fosfát. Jednou důležitou reakcí, která využívá tyto aktivované donory isoprenu, je biosyntéza steroidů. Zde jsou izoprenové jednotky spojeny dohromady za vzniku skvalenu a poté složeny a zformovány do sady prstenců za vzniku lanosterolu. Lanosterol pak může být přeměněn na jiné steroidy, jako je cholesterol a ergosterol.
Odbourávání
Mechanismus odbourávání lipidů se označuje jako β-oxidace. Princip spočívá ve zkracování dlouhého řetězce mastné kyseliny o dva uhlíky. Celý děj β-oxidace probíhá v matrix mitochondrií.
Zdraví
Většina tuků se v potravinách nacházejí ve formě triglyceridů, cholesterolu a fosfolipidů. Některé tuky jsou ve stravě nezbytné, jelikož usnadňují vstřebávání vitamínů, které jsou rozpustné v tucích (A, D, E a K) a karotenoidů.[34]
Lidé a další savci mají dietní požadavky na určité esenciální mastné kyseliny, jako je kyselina linolová (omega-6 mastná kyselina) a alfa-linolenová kyselina (omega-3 mastná kyselina), protože je nelze syntetizovat z jednoduchých prekurzorů ve stravě.[35] Obě tyto mastné kyseliny jsou 18-uhlíkové polynenasycené mastné kyseliny, které se liší v počtu a poloze dvojných vazeb. Většina rostlinných olejů je bohatá na kyselinu linolovou (světlicový, slunečnicový a kukuřičný olej). Kyselina alfa-linolenová se nachází v zelených listech rostlin a v některých semenech, ořeších a luštěninách (zejména lnu, řepce, vlašském ořechu a sóji).[36]
Rybí oleje jsou obzvláště bohaté na omega-3 mastné kyseliny s delším řetězcem, kyselinu eikosapentaenovou (EPA) a kyselinu dokosahexaenovou (DHA). [34]
Mnoho studií prokázalo pozitivní zdravotní přínosy spojené s konzumací omega-3 mastných kyselin u kojenců, rakoviny, kardiovaskulárních chorob a různých duševních chorob (jako deprese, porucha pozornosti s hyperaktivitou a demence).[37][38]
Na rozdíl od toho je nyní dobře známo, že konzumace trans-tuků, které jsou přítomny v částečně hydrogenovaných rostlinných olejích, jsou rizikovým faktorem kardiovaskulárních onemocnění. Tuky, které jsou pro člověka dobré, mohou být přeměněny na trans-tuky nesprávnými způsoby vaření, které vedou k převaření lipidů.[39][40][41]
Několik studií naznačilo, že celkový příjem tuků ve stravě souvisí se zvýšeným rizikem obezity[42][43] a cukrovky,[44] ale řada velmi rozsáhlých studií, včetně studie Women's Health Initiative Dietary Modification Trial, zdravotní studie zdravotních sester a následná studie zdravotníků neodhalila žádné takové vazby.[45][46] Žádná z těchto studií nenaznačila žádnou souvislost mezi procentem kalorií z tuku a rizikem rakoviny, srdečních chorob nebo přibíráním na váze. The Nutrition Source,[47]webová stránka spravovaná oddělením výživy na TH Chan School of Public Health na Harvardově univerzitě, shrnuje aktuální důkazy o vlivu tuků ve stravě: „Podrobný výzkum – z velké části prováděný na Harvardu – ukazuje že celkové množství tuku ve stravě není ve skutečnosti spojeno s váhou nebo nemocí.“[48]
Odkazy
Reference
V tomto článku byly použity překlady textů z článků Lipid na anglické Wikipedii a Lipide na německé Wikipedii.
- ↑ Subramaniam S, Fahy E, Gupta S, Sud M, Byrnes RW, Cotter D, Dinasarapu AR, Maurya MR (October 2011). "Bioinformatics and systems biology of the lipidome". Chemical Reviews. 111 (10): 6452–90. doi:10.1021/cr200295k. PMC 3383319. PMID 21939287.
- ↑ a b c Fahy E, Subramaniam S, Murphy RC, Nishijima M, Raetz CR, Shimizu T, Spener F, van Meer G, Wakelam MJ, Dennis EA (April 2009). "Update of the LIPID MAPS comprehensive classification system for lipids". Journal of Lipid Research. 50 (S1): S9-14. doi:10.1194/jlr.R800095-JLR200. PMC 2674711. PMID 19098281.
- ↑ Mashaghi S, Jadidi T, Koenderink G, Mashaghi A (February 2013). "Lipid nanotechnology". International Journal of Molecular Sciences. 14 (2): 4242–82. doi:10.3390/ijms14024242. PMC 3588097. PMID 23429269.
- ↑ Braconnot H (31 March 1815). "Sur la nature des corps gras". Annales de chimie. 2(XCIII): 225–277.
- ↑ Chevreul ME (1823). Recherches sur les corps gras d'origine animale. Paris: Levrault.
- ↑ Leray C (2012). Introduction to Lipidomics. Boca Raton: CRC Press. ISBN 9781466551466.
- ↑ Leray C (2015). "Introduction, History and Evolution.". Lipids. Nutrition and health. Boca Raton: CRC Press. ISBN 9781482242317.
- ↑ Pelouze TJ, Gélis A (1844). "Mémoire sur l'acide butyrique". Annales de Chimie et de Physique. 10: 434.
- ↑ Comptes rendus hebdomadaires des séances de l'Académie des Sciences, Paris, 1853, 36, 27; Annales de Chimie et de Physique 1854, 41, 216
- ↑ Leray C. "Chronological history of lipid center". Cyberlipid Center. Archived from the original on 2017-10-13. Retrieved 2017-12-01.
- ↑ Prout W (1827). "On the ultimate composition of simple alimentary substances, with some preliminary remarks on the analysis of organised bodies in general". Phil. Trans.: 355–388.
- ↑ Culling CF (1974). "Lipids. (Fats, Lipoids. Lipins).". Handbook of Histopathological Techniques (3rd ed.). London: Butterworths. pp. 351–376. ISBN 9781483164793.
- ↑ Rosenbloom J, Gies WJ (1911). "Suggestion to teachers of biochemistry. I. A proposed chemical classification of lipins, with a note on the intimate relation between cholesterols and bile salts". Biochem. Bull. 1: 51–6.
- ↑ Bloor WR (1920). "Outline of a classication of the lipids". Proc. Soc. Exp. Biol. Med. 17(6): 138–140. doi:10.3181/00379727-17-75. S2CID 75844378.
- ↑ Christie WW, Han X (2010). Lipid Analysis: Isolation, Separation, Identification and Lipidomic Analysis. Bridgwater, England: The Oily Press. ISBN 9780857097866.
- ↑ Bertrand G (1923). "Projet de reforme de la nomenclature de Chimie biologique". Bulletin de la Société de Chimie Biologique. 5: 96–109.
- ↑ Coones RT, Green RJ, Frazier RA (July 2021). "Investigating lipid headgroup composition within epithelial membranes: a systematic review". Soft Matter. 17 (28): 6773–6786. Bibcode:2021SMat...17.6773C. doi:10.1039/D1SM00703C. ISSN 1744-683X. PMID 34212942. S2CID 235708094.
- ↑ Heinz E. (1996). "Plant glycolipids: structure, isolation and analysis", pp. 211–332 in Advances in Lipid Methodology, Vol. 3. W.W. Christie (ed.). Oily Press, Dundee. ISBN 978-0-9514171-6-4
- ↑ Hölzl G, Dörmann P (September 2007). "Structure and function of glycoglycerolipids in plants and bacteria". Progress in Lipid Research. 46(5): 225–43. doi:10.1016/j.plipres.2007.05.001. PMID 17599463.
- ↑ Rosen ED, Spiegelman BM (December 2006). "Adipocytes as regulators of energy balance and glucose homeostasis". Nature. 444(7121): 847–53. Bibcode:2006Natur.444..847R. doi:10.1038/nature05483. PMC 3212857. PMID 17167472.
- ↑ Brasaemle DL (December 2007). "Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis". Journal of Lipid Research. 48(12): 2547–59. doi:10.1194/jlr.R700014-JLR200. PMID 17878492.
- ↑ Malinauskas T, Aricescu AR, Lu W, Siebold C, Jones EY (July 2011). "Modular mechanism of Wnt signaling inhibition by Wnt inhibitory factor 1". Nature Structural & Molecular Biology. 18 (8): 886–93. doi:10.1038/nsmb.2081. PMC 3430870. PMID 21743455.
- ↑ Malinauskas T (March 2008). "Docking of fatty acids into the WIF domain of the human Wnt inhibitory factor-1". Lipids. 43 (3): 227–30. doi:10.1007/s11745-007-3144-3. PMID 18256869. S2CID 31357937.
- ↑ Wang X (June 2004). "Lipid signaling". Current Opinion in Plant Biology. 7 (3): 329–36. doi:10.1016/j.pbi.2004.03.012. PMID 15134755.
- ↑ Dinasarapu AR, Saunders B, Ozerlat I, Azam K, Subramaniam S (June 2011). "Signaling gateway molecule pages--a data model perspective". Bioinformatics. 27 (12): 1736–8. doi:10.1093/bioinformatics/btr190. PMC 3106186. PMID 21505029.
- ↑ Indiveri C, Tonazzi A, Palmieri F (October 1991). "Characterization of the unidirectional transport of carnitine catalyzed by the reconstituted carnitine carrier from rat liver mitochondria". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1069 (1): 110–6. doi:10.1016/0005-2736(91)90110-t. PMID 1932043.
- ↑ Parodi AJ, Leloir LF (April 1979). "The role of lipid intermediates in the glycosylation of proteins in the eucaryotic cell". Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes. 559 (1): 1–37. doi:10.1016/0304-4157(79)90006-6. PMID 375981.
- ↑ Helenius A, Aebi M (March 2001). "Intracellular functions of N-linked glycans". Science. 291 (5512): 2364–9. Bibcode:2001Sci...291.2364H. doi:10.1126/science.291.5512.2364. PMID 11269317. S2CID 7277949.
- ↑ Nowicki M, Müller F, Frentzen M (April 2005). "Cardiolipin synthase of Arabidopsis thaliana". FEBS Letters. 579 (10): 2161–5. doi:10.1016/j.febslet.2005.03.007. PMID 15811335. S2CID 21937549.
- ↑ Gohil VM, Greenberg ML (February 2009). "Mitochondrial membrane biogenesis: phospholipids and proteins go hand in hand". The Journal of Cell Biology. 184 (4): 469–72. doi:10.1083/jcb.200901127. PMC 2654137. PMID 19237595.
- ↑ Hoch FL (March 1992). "Cardiolipins and biomembrane function"(PDF). Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes. 1113 (1): 71–133. doi:10.1016/0304-4157(92)90035-9. hdl:2027.42/30145. PMID 1550861.
- ↑ "Steroids". Elmhurst. edu. Archived from the original on 2011-10-23. Retrieved 2013-10-10.
- ↑ Stryer L, Berg JM, Tymoczko JL (2007). Biochemistry (6th ed.). San Francisco: W.H. Freeman. ISBN 978-0-7167-8724-2.
- ↑ a b Bhagavan NV (2002). Medical Biochemistry. San Diego: Harcourt/Academic Press. ISBN 978-0-12-095440-7.
- ↑ Stryer L, Berg JM, Tymoczko JL (2007). Biochemistry(6th ed.). San Francisco: W.H. Freeman. ISBN 978-0-7167-8724-2.
- ↑ Russo GL (March 2009). "Dietary n-6 and n-3 polyunsaturated fatty acids: from biochemistry to clinical implications in cardiovascular prevention". Biochemical Pharmacology. 77 (6): 937–46. doi:10.1016/j.bcp.2008.10.020. PMID 19022225.
- ↑ Riediger ND, Othman RA, Suh M, Moghadasian MH (April 2009). "A systemic review of the roles of n-3 fatty acids in health and disease". Journal of the American Dietetic Association. 109 (4): 668–79. doi:10.1016/j.jada.2008.12.022. PMID 19328262.
- ↑ Galli C, Risé P (2009). "Fish consumption, omega 3 fatty acids and cardiovascular disease. The science and the clinical trials". Nutrition and Health. 20 (1): 11–20. doi:10.1177/026010600902000102. PMID 19326716. S2CID 20742062.
- ↑ Micha R, Mozaffarian D (2008). "Trans fatty acids: effects on cardiometabolic health and implications for policy". Prostaglandins, Leukotrienes, and Essential Fatty Acids. 79 (3–5): 147–52. doi:10.1016/j.plefa.2008.09.008. PMC 2639783. PMID 18996687.
- ↑ Dalainas I, Ioannou HP (April 2008). "The role of trans fatty acids in atherosclerosis, cardiovascular disease and infant development". International Angiology. 27 (2): 146–56. PMID 18427401.
- ↑ Mozaffarian D, Willett WC (December 2007). "Trans fatty acids and cardiovascular risk: a unique cardiometabolic imprint?". Current Atherosclerosis Reports. 9 (6): 486–93. doi:10.1007/s11883-007-0065-9. PMID 18377789. S2CID 24998042.
- ↑ Astrup A, Dyerberg J, Selleck M, Stender S (2008), "Nutrition transition and its relationship to the development of obesity and related chronic diseases", Obes Rev, 9 (S1): 48–52, doi:10.1111/j.1467-789X.2007.00438.x, PMID 18307699, S2CID 34030743
- ↑ Astrup A (February 2005). "The role of dietary fat in obesity". Seminars in Vascular Medicine. 5 (1): 40–7. doi:10.1055/s-2005-871740. PMID 15968579.
- ↑ Astrup A (2008). "Dietary management of obesity". Journal of Parenteral and Enteral Nutrition. 32 (5): 575–7. doi:10.1177/0148607108321707. PMID 18753397.
- ↑ Beresford SA, Johnson KC, Ritenbaugh C, Lasser NL, Snetselaar LG, Black HR, et al. (February 2006). "Low-fat dietary pattern and risk of colorectal cancer: the Women's Health Initiative Randomized Controlled Dietary Modification Trial". Journal of the American Medical Association. 295 (6): 643–54. doi:10.1001/jama.295.6.643. PMID 16467233.
- ↑ Howard BV, Manson JE, Stefanick ML, Beresford SA, Frank G, Jones B, Rodabough RJ, Snetselaar L, Thomson C, Tinker L, Vitolins M, Prentice R (January 2006). "Low-fat dietary pattern and weight change over 7 years: the Women's Health Initiative Dietary Modification Trial". Journal of the American Medical Association. 295 (1): 39–49. doi:10.1001/jama.295.1.39. PMID 16391215.
- ↑ "The Nutrition Source". T. H. Chan School of Public Health. Harvard University.
- ↑ "Fats and Cholesterol: Out with the Bad, In with the Good — What Should You Eat? – The Nutrition Source". Harvard School of Public Health.
Související články
Externí odkazy
- Obrázky, zvuky či videa k tématu lipid na Wikimedia Commons
- Slovníkové heslo lipid ve Wikislovníku
- Lipidy na webu Biotox
