Carcharocles megalodon
|
| |
|---|---|
 Model čelistí v Americkém přírodovědném muzeu
| |
| Vědecká klasifikace | |
| Říše |
živočichové (Animalia) |
| Kmen |
strunatci (Chordata) |
| Třída |
paryby (Chondichthyes) |
| Nadřád |
žraloci (Selachimoha) |
| Řád |
obrouni (Lamnifomes) |
| Čeleď |
Otodontidae |
| Rod |
Carcharocles |
| Binomické jméno | |
|
Carcharocles megalodon (Agassiz, 1843) | |
| Některá data mohou pocházet z datové položky. | |
Megalodon (Carcharocles megalodon, v překladu „velký zub“, nebo také Otodus megalodon) je vyhynulý druh žraloka, který žil před asi 23 až 3,6 milionu let během období od raného miocénu do pliocénu. O systematice tohoto tvora probíhaly debaty. Dle jedněch názorů patřil do čeledi lamnovití (Lamnidae) a byl blízkým příbuzným velkého žraloka bílého (Carcharodon carcharias), dle druhých do vyhynulé čeledi Otodontidae; tento předpoklad je pravděpodobně správný. Diskutuje se rovněž o umístění v rodu Carcharocles, Megaselachus, Otodus nebo Procarcharodon.
Megalodon mohl vzhledem připomínat mohutnějšího žraloka bílého či se mohl podobat žraloku velikému (Cetorhinus maximus) či písečnému (Carcharias taurus). Fosilní pozůstatky naznačují, že tento žralok mohl měřit na délku až v rozmezí 18 až 20 metrů. Tyto obří rozměry se u něho objevily patrně z důvodu postupného evolučního vývoje v ochlazujících se vodách.[1] Jeho velké čelisti, vybavené silnými a robustními zuby sloužícími k zabíjení a trhání kořisti, mohly vyvinout při skousnutí sílu 108 500 až 182 200 newtonů.
Megalodoni měli asi silný vliv na strukturu mořských společenstev. Na základě fosilií se zdá, že se jednalo o kosmopolita, tedy druh žijící ve všech oceánech. Zřejmě se zaměřoval na velkou kořist, jako byly velryby, ploutvonožci a velké druhy želv té doby. Na rozdíl od žraloka bílého, který na kořist útočí zespodu, megalodon pravděpodobně využíval své silné čelisti k prokousnutí hrudní dutiny oběti a poškození vnitřních orgánů – srdce a plic. Některé výzkumy naznačují, že tento obří žralok mohl být endotermní (udržoval si stálou tělesnou teplotu).[2]
Megalodon byl v kompetičním vztahu s kytovci lovícími jiné velryby, kteří možná přispěli k jeho vyhynutí. Vzhledem k tomu, že žil primárně v teplých vodách, za jeho vyhynutí mohl i nástup ledové doby či ochlazení oceánů. Zároveň poklesla hladina moře, čímž žraloci přišli o území, kde vyrůstala jejich mláďata. Zmenšil se rovněž počet druhů kosticovců, kteří se zároveň začali přesouvat do polárních oblastí; tím megalodon přišel o hlavní zdroj potravy. Vyhynutí tohoto velkého žraloka ovlivnilo ostatní mořské tvory; poté, co vymizel, velikost kosticovců výrazně vzrostla.
Taxonomie
Pojmenování
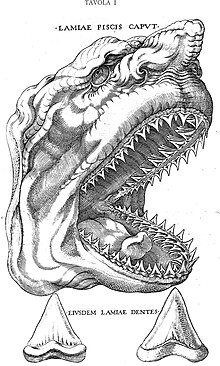
V době renesance byly obrovské trojúhelníkové fosilní zuby, nalézané převážně ve skalních útvarech, považovány za zkamenělé jazyky draků a hadů a nazývány jako glossopetrae. Teprve v roce 1667 tento názor vyvrátil dánský přírodovědec Nicolas Steno, který je správně popsal jako zuby žraloka, a vytvořil rovněž realistický nákres hlavy žraloka, který takové zuby mohl vlastnit. Své poznatky popsal ve své monografii De glossopetris dissertatio, která obsahovala i ilustraci tohoto zubu.[3][4][5]
Poprvé tento druh popsal švýcarský přírodovědec Louis Agassiz pod vědeckým jménem Carcharodon megalodon. Do rodu Carcharodon jej zařadil vzhledem k podobnosti zubů se zuby žraloka bílého (Carcharodon carcharias) ze stejného rodu. Jeho popis, založený na zbytcích zubů, uvedl ve své práci Recherches sur les poissons fossiles z roku 1843.[6][7]Anglický paleontolog Edward Charlesworth však ve své práci již z roku 1837 určil pro tento druh název Carcharias megalodon, přičemž v ní citoval Agassize. Z toho plyne, že Agassiz ve skutečnosti megalodona popsal ještě před rokem 1843, pravděpodobně ve svých předchozích článcích z roku 1835.[8]
Název megalodon v překladu znamená „velký zub“, jméno pochází ze dvou starořeckých slov: μέγας (mégas), znamenající velký či mocný, a ὀδούς (odoús), což znamená zub.[9][10] Toto pojmenování je nicméně pouze neformální.[11]
Evoluce
Nejstarší pozůstatky megalodona pocházejí z pozdního oligocénu před zhruba 28 miliony lety,[12][13] přesto existují i další názory, že se druh objevil například před 16 nebo 23 miliony lety. Megalodon údajně vyhynul během konce pliocénu před asi 2,6 miliony lety,[14][15] výzkum z roku 2019 zasadil vyhynutí megalodona do doby před 3,6 miliony lety.[16] Existují prý i záznamy o zubech z následujícího období pleistocén, ale jsou považovány za pochybné.[15]

Megalodon je považován za zástupce čeledi Otodontidae, v níž tvoří část rodu Carcharocles; na základě předchozí systematiky byl nicméně řazen mezi lamnovité (Lamnidae), jako zástupce rodu Carcharodon.[14][15][17][18]
Do rodu Carcharodon byl zařazen na základě podobnosti zubů se žralokem bílým, je však pravděpodobnější, že se obě paryby vyvíjely konvergentně, tedy pod stejnými selekčními tlaky, díky čemuž se vyvinul podobný vzhled obou zvířat, přestože společně nemusela býti blízce příbuzná. Žralok bílý je naopak považován spíše za příbuzného žraloka druhu Isurus hastalis (zuby těchto dvou druhů jsou podobnější, megalodon měl jemnější vroubkování). Společný předek žraloka bílého a rodu Isurus žil před asi 4 miliony lety.[7][19] Naopak dle teorie příbuznosti megalodona a žraloka bílého jsou prý rozdíly v zubech nepatrné a nejasné.[20]

Rod Carcharocles je zastoupen čtyřmi druhy: Carcharocles auriculatus, Carcharocles angustidens, Carcharocles chubutensis a Carcharocles megalodon.[21] Určili jej v roce 1923 výzkumníci D. S. Jordan a H. Hannibal a prvotně obsahoval pouze druh C. auriculatus, megalodon sem byl zařazen až v 80. letech 20. století.[7][21] Předtím, v roce 1960, francouzský ichtyolog Edgard Casier vytvořil rod Procarcharodon, kam zařadil tyto čtyři druhy, jež považoval za nepříbuzné se žralokem bílým; tento rod se stal mladším synonymem rodu Carcharocles.[21] Vedle rodu Procarcharodon byl postaven rod Palaeocarcharodon, který měl být začátkem vývojové linie. Zastánci teorie o příbuznosti žraloka bílého a megalodona potom považují Paleocarcharodon za posledního společného předka těchto dvou druhů, dle odpůrců teorie se jedná o slepou vývojovou větev.[22]
Existuje i další model evoluce megalodona, který také navrhl Casier ve stejném roce. Podle tohoto názoru je předkem rodu Carcharocles žralok Otodus obliquus, který žil během paleocénu až miocénu.[19][23] Ten se podle této teorie vyvinul na druh Otodus aksuaticus a vývoj dále pokračoval skrze druhy Carcharocles auriculatus, Carcharocles angustidens a Carcharocles chubutensis až po megalodona. Vývoj této linie je charakterizován postupným narůstáním vroubkování zubů, rozšiřováním jejich korunky, nabýváním trojúhelníkového tvaru a ztrátou bočních hrotů.[24][23] Předky rodu Otodus mají být žraloci Cretolamna z období křídy.[17][25]
Další model evoluce rodu Carcharocles představil v roce 2001 paleontolog Michael Benton. Druhy Carcharocles auriculatus, Carcharocles angustidens a Carcharocles chubutensis jsou dle něj ve skutečnosti jediným druhem, který se v průběhu času jenom postupně měnil.[26][13][27] Na základě dalších studií by Carcharocles auriculatus, Carcharocles angustidens a Carcharocles chubutensis měly tvořit jediný druh rodu Otodus, megalodon by pak byl jediným zástupcem rodu Carcharocles.[13][28]
Další možností je zneplatnění rodu Carcharocles a řazení megalodona přímo do rodu Otodus, jako Otodus megalodon.[pozn. 1] Objev fosilií přiřazených rodu Megalolamna v roce 2016 vedl k dalším zkoumáním rodu Otodus, přičemž se zdá se, že rod je parafyletický, tedy zahrnuje posledního společného předka, ale ne všechny jeho potomky. Pokud by byl do tohoto rodu zařazen současný rod Carcharocles, byla by zřejmě vytyčena celá fylogenetická linie (monofyletismus). Rod Megalolamna by se v tomto případě stal sesterskou linií.[17]
Biologie
Vzhled

Existuje více možných teorií popisujících vzhled megalodona. Na základě jedné se jednalo o mohutného žraloka podobného žralokovi bílému (Carcharodon carcharias), avšak s většími a širšími čelistmi než má on. Ploutve byly dle této teorie u megalodona podobné jako u žraloka bílého, ale silnější. Jeho hluboké drobné oči mohly připomínat prasečí.[30]
Další teorií je, že megalodon připomínal žraloka obrovského (Rhincodon typus) či velikého (Cetorhinus maximus). V tomto případě by asi ocasní ploutev měla tvar půlměsíce a žralok by byl vybaven malou řitní a druhou hřbetní ploutví. Po obou stranách ocasu se v tomto případě mohl vyvinout vodorovný kýl. Ten se hojně vyskytuje i u jiných velkých vodních živočichů, jako jsou velryby, tuňáci a další žraloci a usnadňuje plavání. Tvar hlavy však nelze odhadnout, protože se mezi jednotlivými druhy liší; většina tělesných adaptací, majících snižovat odpor vody, se vyvíjela spíše směrem k ocasu.[31]

Vzhledem k tomu, že rod Carcharocles se pravděpodobně vyvinul z rodu Otodus, a oba mají zuby podobné žralokům písečným (Carcharias taurus), dle nepravděpodobné teorie je možné i to, že megalodon vypadal podobně jako tento žralok.[31][32]
Rozměry
![Porovnání megalodona (šedý=největší odhadovaná velikost, červený=nejmenší odhadovaná velikost) se žralokem velrybím (fialově), bílým (červeně) a člověkem, ale megalodon mohl mít štíhlejší a protáhlejší tělo.[33]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/4d/Megalodon_scale.svg/220px-Megalodon_scale.svg.png)
Vzhledem k tomu, že se z megalodona dochovaly pouze zuby a obratle, existuje mnoho protikladných odhadů jeho velikosti.[34][35] I kvůli nedostatku fosilního materiálu jsou jeho popis a velikost odhadovány na základě žraloka bílého,[36] který je považován za tvora jemu nejpodobnějšího.[28]
Roku 1909 americký zoolog Bashford Dean poprvé rekonstruoval čelisti megalodona, které jsou součástí sbírek Amerického přírodovědného muzea. Z rozměrů této rekonstrukce se předpokládalo, že megalodon měřil na délku asi 30 metrů. Ve své rekonstrukci však Dean přecenil velikost čelistí, což způsobilo tento přehnaný údaj.[37][38]
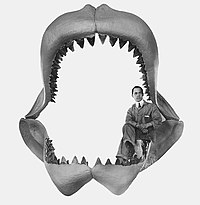

Ichtyolog John E. Randall v 70. letech 20. století využil k odhadnutí velikosti megalodona výšku zubní skloviny. Na základě ní mu vyšlo, že žralok měřil okolo třinácti metrů.[39] Výška skloviny se nicméně nemusí zvyšovat v přesném poměru k celkové délce zvířete.[40]
V 90. letech 20. století dva mořští biologové Patrick J. Schembri a Stephen Papson určili megalodonovu velikost až na 24–25 metrů.[41][42]
Odborníci na žraloky Michael D. Gottfried, Leonard Compagno a S. Curtis Bowman navrhli jiný postup, a sice lineární vztah mezi celkovou délkou žraloka a výškou největšího předního zubu.[13][43] Na základě výšky největšího objeveného zubu odhadli průměrnou délku velkých samic megalodona na 15,6 metrů, s tím, že existovat mohou i větší zuby.[44]
Roku 2002 přišel s dalším návrhem výzkumník žraloků Clifford Jeremiah. Podle něj byla celková délka přímo úměrná s šířkou kořenu horního předního zubu. Na základě této teorie měl 1 cm šířky kořenu odpovídat přibližně 1,4 m délky žraloka. Jeremiah svou teorii podporoval tím, že obvod vnějších čelistí žraloků je přímo uměrný celkové délce paryby, přičemž pomocí šířky kořenů zubů se určuje přibližná délka obvodu čelistí. Tímto způsobem Jeremiah na základě nejdelšího megalodonova zubu (šířka kořene 12 cm), který vlastnil, odhadnul délku megalodona na 16,5 metru.[45]
Roku 2002 paleontolog Kenshu Shimada z DePaulovy univerzity navrhl lineární vztah mezi výškou zubní korunky a celkovou délkou. Na základě anatomické analýzy několika vzorků byl tento vztah univerzální pro kterýkoli zub. Na základě tohoto modelu určil pro Gottfriedův zub velikost žraloka asi 15 metrů.[46]
Na základě novějších studií z let 2014–2017 byla určena velikost maximálně 18 metrů s průměrem 10,5 metru[14][15][18] (největší žraloci bílí měřili 6,1 metru). Je možné, že různé populace megalodonů dosahovaly na různých územích odlišných velikostí kvůli různým ekologickým tlakům.[18] Pokud by žralok dosahoval větší velikosti než 16 metrů, byla by to největší známá (pa)ryba, která kdy žila, a překonal by i obrovskou jurskou rybu Leedsichthys.[37]
Studie publikovaná v říjnu roku 2019 přehodnocuje dřívější odhady velikosti megalodona (délka 18 až 20 metrů) jako mírně nadhodnocené a na základě výzkumu zubů stanovuje jako pravděpodobnou délku dospělých exemplářů megalodona 14,2 až 15,3 metru. Exempláře dlouhé přes 15 metrů lze přitom považovat za extrémně vzácné, přesto zůstává tento žralok jedním z největších známých predátorů v dějinách života na Zemi.[47][48] Podle výsledků odborné studie z roku 2020 byl megalodon dlouhý asi 16 metrů, jeho hřbetní ploutev byla vysoká zhruba 1,62 metru a jeho ocasní ploutev měřila na výšku celkově 3,85 metru. Jeho hlava pak byla dlouhá asi 4,65 metru, tedy zhruba jako delší automobil.[49][50] Jiné odhady z roku 2021 opět udávají maximální délku dospělců tohoto druhu na rovných 20 metrů.[51] Megalodon byl s délkou minimálně 14 metrů skutečně výrazně nad velikostním limitem pro všechny ostatní lamniformní žraloky, u nichž délka nepřesahovala přibližně 7 metrů.[52]
Dospělí samci megalodona mohli vážit mezi 12,6 až 33,9 tun a samice mezi 27,4 a 59,4 tun. Samice byly celkově větší, o velikosti asi 13,3 až 17 m, kdežto samci měřili mezi 10,5 až 14,3 m.[53][54] Na základě studie vydané roku 2015, která zkoumala rychlost žraloků na základě jejich velikosti, bylo odhadnuto, že megalodon plaval rychlostí asi 18 kilometrů za hodinu, což odpovídá jeho průměrné hmotnosti 48 tun. Výsledek odpovídal rychlosti jiných vodních tvorů o podobné velikosti: například podobně velký plejtvák myšok (Balaenoptera physalus) se pohybuje rychlostí mezi 14,5–21,5 km/h.[55]
Obří velikost megalodona mohla být způsobena klimatickými faktory, množstvím velké kořisti či vývojem mezotermie – přechodem mezi teplokrevností a studenokrevností, která by vedla ke zvýšení rychlosti metabolismu megalodona a ten by pak dovedl také rychleji plavat. Nicméně, protože jsou žraloci z čeledi Otodontidae považováni za studenokrevné a megalodon je považován za jejich blízkého příbuzného, je možné, že i on byl studenokrevný. Tato možnost je ale nepravděpodobná, vzhledem k tomu, že se většina současných studenokrevných žraloků, jako například žralok obrovský, živí převážně filtrováním potravy z vody, a tedy nejsou adaptováni na dravý styl života.[56]
Zuby a síla skusu

Nejčastěji zachované fosilie megalodona jsou zuby. Ty jsou velké a robustní, mají trojúhelníkový tvar s napojením kořene na korunku ve tvaru písmene V, malé vroubkování a postrádají postranní hroty.[28][57] Zub byl v čelisti podobně jako u žraloka bílého zasazen strmě. Zuby byly v čelisti ukotveny vlákny kolagenu, jejich základna byla drsná, což přidávalo na pevnosti.[58] Strana zubu směřující k jazyku byla vypouklá (konvexní), druhá strana zubu pouze mírně konvexní až skoro plochá. Přední zuby byly téměř kolmé k čelisti a symetrické, zadní zasazeny šikmo a asymetrické.[59]
Zuby megalodona mohou měřit ve výšce sklonu (diagonální délka) i přes 180 milimetrů.[60] V rekonstrukci čelistí od lovce fosilií Vita Bertucciho je obsažen i zub o údajné délce více než 180 mm.[61][62] S těmito rozměry jsou zuby megalodona největšími mezi všemi doposud známými druhy žraloků.[60] Roku 1989 byla v japonské Saitamě objevena jejich téměř kompletní sada, další sada zubů megalodona byla vykopána v americké formaci Yorktown (Maryland) a posloužila jako základ pro rekonstrukci čelistí v Národním přírodovědeckém muzeu ve Washingtonu. Na základě těchto objevů byl v roce 1996 určen celkový zubní vzorec megalodona.[57][63]

Ten činí 2.1.7.43.0.8.4. Jak je patrné ze vzorce, megalodon měl v čelistech čtyři druhy zubů: přední, střední, boční a zadní. Měl velmi robustní chrup,[64] čelisti byly osazeny až 250 zuby v pěti řadách.[65] Zoubkování jejich nositeli usnadnilo prořezávání masem a kostmi. Čelist u velkých jedinců měřila napříč až dva metry a mohla se rozevřít až do 75°, rekonstrukce v Národním přírodovědeckém muzeu se blíží i 100°.[7][66][67]
V roce 2008 uskutečnil tým vědců pod vedením S. Wroea pokus mající zjistit sílu skusu velkého bílého žraloka. Pokus byl proveden s 2,5metrovým exemplářem a výsledky poté přepočteny na maximální potvrzenou velikost žraloka a rovněž na minimální a maximální tělesnou hmotnost megalodona. Podle výsledků mohl megalodon vyvinout sílu o 108 514 až 182 201 newtonech, ve srovnání s 18 216 newtony u žraloka bílého a 5 300 až 7 400 u vyhynulé ryby Dunkleosteus. Kromě toho tým uvedl, že žraloci sebou při útoku na kořist trhají, což zvyšuje udanou sílu, čímž bude pravděpodobně celková působená síla ještě vyšší než odhad.[54][68][69]
Vnitřní popis

Fosilie megalodona jsou zastoupeny pouze zuby, obratli a koprolity – fosilním trusem.[36] Stejně jako u všech žraloků, i kostra megalodona nebyla tvořena kostmi, ale spíše rychle se rozpadající chrupavkou; proto je většina fosilních vzorků špatně zachována.[70][71] K podpoře jeho mohutných zubů by musely být čelisti megalodona mnohem masivnější, pevnější a silnější než čelisti žraloka bílého. Rovněž jeho chrupavčitá lebka, chondrokranium, by byla mohutnější než u žraloka bílého, i ploutve by se u megalodona vyvinuly velké.[30]
Fosilní obratle megalodona byly nalezeny například v oblasti Antverp v Belgii. Tento pozoruhodný nález obsahoval 150 částečně zachovaných obratlů, měřících mezi 55 až 155 milimetry v průměru (ovšem obratle megalodona mohly být i mnohem větší). Další výzkum tohoto vzorku ukázal, že megalodon měl nejvyšší počet obratlů mezi všemi žraloky, možná přes 200 obratlů; podobnému počtu se přibližuje pouze žralok bílý.[72] Další podobně zachovalé obratle byly vyzvednuty v 80. letech 20. století v Dánsku ve formaci Gram. Bylo objeveno celkem dvacet obratlů, o délce mezi 100 až 230 mm.[58]

Koprolity megalodona byly nalezeny například v Jižní Karolíně a datovány do období miocén. Měřily 14 cm a byly spirálovité, což naznačuje, že byl megalodon vybaven takzvanou spirální řasou, spodní stočenou částí střev, která pomáhá lepší absorpci živin. Tuto adaptaci mají například současní lamnovití.[73]
Kompletní rekonstrukce kostry megalodona, která byla vystavována v americkém Calvert Marine Museum a jihoafrickém Iziko South African Museum, byla vytvořena vědcem Michaelem Gottfriedem a kolektivem.[74] Tento 11,3 metrů dlouhý exponát má představovat dospělého samce.[53]
Paleobiologie
Výskyt
Megalodon byl kosmopolitem;[14][75] jeho fosilie byly vykopány na mnoha místech světa, včetně Evropy, Afriky, Ameriky a Austrálie.[76][77] Nejčastěji se vyskytoval v subtropických až mírných zeměpisných šířkách[14][78] až do 55° severní šířky. Toleroval vody o teplotách mezi 1–24 °C. Je pravděpodobné, že podobně nízké teploty snášel díky mezotermii, fyziologické schopnosti velkých žraloků, pomocí které dovedou udržovat vyšší teplotu těla než je teplota vody[14] (výroba metabolického tepla zaznamenaná právě například u žraloků).
Megalodon obýval širokou škálu mořských oblastí, jako jsou mělké pobřežní vody, bažinaté laguny, písčitý litorál i hluboké moře a vykazoval přechodný životní styl. Dospělí megalodoni nebyli hojní v mělkých vodách, většinou obývali pobřežní oblasti. Zejména v různých fázích životního cyklu se mohl megalodon pohybovat mezi pobřežními a vzdálenějšími oceánskými vodami.[60][79] Jeho fosilie byly pak objeveny i tak daleko od pevniny, jako u Marianského příkopu v Tichém oceánu.[65]
Fosilní pozůstatky naznačují, že megalodoni žijící na jižní polokouli byli v průměru větší než megalodoni ze severní polokoule. Jedinci žijící v Tichém oceánu potom byli větší než megalodoni v oceánu Atlantském. Není však předpokládán žádný trend zvětšování velikosti těla v určité zeměpisné šířce či změna velikosti v průběhu času (přestože linie Carcharocles tuto vlastnost vykazuje). Modus všech velikostí byl odhadnut na asi 10,5 m, přičemž větší jedinci obývají více oblastí. To naznačuje ekologickou nebo konkurenční výhodu pro větší velikost těla.[18]
Rozmístění fosilií
Megalodon se vyskytoval po celém světě a objeveno bylo mnoho fosilií ve všech oceánech neogénu.[80]
| Věk | souvrství | Stát | Kontinent |
|---|---|---|---|
| pliocén | souvrství Luanda |
|
Afrika |
|
|
Afrika | ||
|
|
Afrika | ||
| souvrství Castell'Arquato |
|
Evropa | |
| souvrství Arenas de Huelva |
|
Evropa | |
| souvrství Esbarrondadoiro |
|
Evropa | |
| souvrství Touril Complex |
|
Evropa | |
| souvrství Red Crag |
|
Evropa | |
| souvrství San Mateo |
|
Severní Amerika | |
| souvrství Towsley |
|
Severní Amerika | |
| souvrství Bone Valley |
|
Severní Amerika | |
| souvrství Tamiami |
|
Severní Amerika | |
| souvrství Yorktown |
|
Severní Amerika | |
| souvrství Highlands |
|
Severní Amerika | |
| souvrství Refugio |
|
Severní Amerika | |
| souvrství San Diego |
|
Severní Amerika | |
| souvrství Tirabuzon |
|
Severní Amerika | |
| souvrství Onzole |
|
Jižní Amerika | |
| souvrství Paraguaná |
|
Jižní Amerika | |
| pískovce Black Rock |
|
Oceánie | |
| souvrství Cameron Inlet |
|
Oceánie | |
| souvrství Grange Burn |
|
Oceánie | |
| souvrství Loxton Sand |
|
Oceánie | |
| souvrství Whaler's Bluff |
|
Oceánie | |
| souvrství Tangahoe |
|
Oceánie | |
| miocén | |||
|
|
Afrika | ||
| Madagaskarská pánev |
|
Afrika | |
|
|
Afrika | ||
| souvrství Varswater |
|
Afrika | |
| vápence Baripada |
|
Asie | |
| souvrství Arakida |
|
Asie | |
| Bihoku |
|
Asie | |
| souvrství Fudžina |
|
Asie | |
| souvrství Hannoura |
|
Asie | |
| souvrství Hongo |
|
Asie | |
| souvrství Horimacu |
|
Asie | |
| souvrství Ičiši |
|
Asie | |
| souvrství Kurahara |
|
Asie | |
| souvrství Maenami |
|
Asie | |
| Macujama |
|
Asie | |
| souvrství Sekinobana |
|
Asie | |
| souvrství Suso |
|
Asie | |
| souvrství Takakubo |
|
Asie | |
| souvrství Tonokita |
|
Asie | |
| souvrství Curuši |
|
Asie | |
| souvrství Wadžimazaki |
|
Asie | |
| souvrství Jošii |
|
Asie | |
|
|
Asie | ||
| souvrství Burgeschleinitz |
|
Evropa | |
| souvrství Melker Sand |
|
Evropa | |
| souvrství Rzehakia |
|
Evropa | |
| souvrství Weissenegg |
|
Evropa | |
| Antwerpen Sands Member |
|
Evropa | |
|
|
Evropa | ||
| souvrství Hrušky |
|
Evropa | |
| souvrství Gram |
|
Evropa | |
| Akvitánská pánev |
|
Evropa | |
|
|
Evropa | ||
| pískovce Libano |
|
Evropa | |
| souvrství Blue Clay |
|
Evropa | |
| vápence Globigerina |
|
Evropa | |
| Aalten Member |
|
Evropa | |
| souvrství Breda |
|
Evropa | |
| Korytnické jíly |
|
Evropa | |
| vápence Leitha |
|
Evropa | |
| souvrství Esbarrondadoiro |
|
Evropa | |
| souvrství Fiľakovo |
|
Evropa | |
| souvrství Arjona |
|
Evropa | |
| kalcarenity Sant Elm |
|
Evropa | |
|
|
Evropa | ||
| souvrství Monterey |
|
Severní Amerika | |
| souvrství Puente |
|
Severní Amerika | |
| souvrství Purisima |
|
Severní Amerika | |
| souvrství San Mateo |
|
Severní Amerika | |
| souvrství Santa Margarita |
|
Severní Amerika | |
| souvrství Temblor |
|
Severní Amerika | |
| souvrství Topanga |
|
Severní Amerika | |
| souvrství Bone Valley |
|
Severní Amerika | |
| souvrství Calvert |
|
Severní Amerika | |
| souvrství Kirkwood |
|
Severní Amerika | |
|
|
Severní Amerika | ||
| souvrství Cojímar |
|
Severní Amerika | |
| souvrství Kendance |
|
Severní Amerika | |
|
|
Severní Amerika | ||
| vápence Aymamón |
|
Severní Amerika | |
| souvrství Almejas |
|
Severní Amerika | |
| souvrství Carrillo Puerto |
|
Severní Amerika | |
| souvrství Chagres |
|
Severní Amerika | |
| souvrství Chucunaque |
|
Severní Amerika | |
| souvrství Gatun |
|
Severní Amerika | |
| souvrství Paraná |
|
Jižní Amerika | |
| souvrství Bahía Inglesa |
|
Jižní Amerika | |
| souvrství Castilletes |
|
Jižní Amerika | |
| souvrství Miramar |
|
Jižní Amerika | |
| souvrství Pisco |
|
Jižní Amerika | |
| souvrství Camacho |
|
Jižní Amerika | |
| souvrství Cantaure |
|
Jižní Amerika | |
| souvrství Caujarao |
|
Jižní Amerika | |
| souvrství Socorro |
|
Jižní Amerika | |
| souvrství Urumaco |
|
Jižní Amerika | |
| vápence Batesford |
|
Oceánie | |
| pískovce Black Rock |
|
Oceánie | |
| vápence Gippsland |
|
Oceánie | |
| souvrství Mannum |
|
Oceánie | |
| vápence Morgan |
|
Oceánie | |
| vápence Port Campbell |
|
Oceánie | |
|
|
Oceánie | ||
|
|
Oceánie |
Potrava

Žraloci se obecně živí oportunisticky, ale je možné, že megalodon byl obzvláště dravý. Jeho velká velikost společně se schopností rychlého plavání a mocnými čelistmi z něj činila vrcholového predátora schopného zabíjet široké spektrum kořisti.[54][81] Studie zkoumající izotopy vápníku vyhynulých a žijících paryb ukázala, že megalodon byl na vyšší trofické úrovni než žralok bílý; byl tedy v potravním řetězci ještě výše.[82]
Fosilní důkazy naznačují, že se tento druh živil mnoha druhy kytovců, jako jsou delfíni, malé velryby, velryby z čeledi cetotheridů či squalodontidů, dále zabíjel vorvaně, velryby grónské či plejtvákovité.[37][83][84] Mimo to se živil sirénami, velkými mořskými želvami,[79] nepohrdl ani menšími rybami a dalšími druhy žraloků.[37][71] Bylo objeveno mnoho velrybích kostí s hlubokými kousanci pravděpodobně od megalodonů, jejichž zuby se navíc někdy našly blízko nebo přímo ve velrybích ostatcích.[11][23][85]
Zdá se, že složení potravy megalodonů se měnilo s věkem a stanovišti, podobně jako u současných žraloků bílých. Například megalodoni žijící při pobřeží dnešního státu Peru se pravděpodobně zaměřovali na lov 2,5 až 7metrových cetotheridů a jiné kořisti menší než byli oni sami, větší druhy velryb zabíjeli méně.[83] Mladí jedinci pravděpodobně konzumovali spíše ryby.[28][86]
Kompetiční vztahy

Megalodon žil ve vysoce konkurenčním prostředí.[87] Nicméně jeho postavení na vrcholu potravního řetězce[88] mělo pravděpodobně velký vliv na strukturu mořských společenstev.[87][89] Fosilní důkazy naznačují vzájemný vztah mezi existencí tohoto mořského predátora a rozvojem kytovců a dalších velkých mořských savců.[78][87] Mladí megalodoni pravděpodobně preferovali stanoviště s dostatkem malých druhů kytovců, kdežto dospělci vyhledávali oblasti s dostatkem kytovců velkých. Tyto preference se mohly vyvinout krátce poté, co se megalodon poprvé objevil v oligocénu.[90]
Megalodon koexistoval společně s ozubenými kytovci (především squalodontidé a vorvani), kteří byli pravděpodobně také vrcholovými predátory, a byli tedy s megalodonem v kompetičním vztahu.[87] Konkurence odstartovala u ozubených evoluční závody ve zbrojení. Některé druhy, jako například kosatky Orcinus citoniensis, začaly lovit ve skupinách,[23][91] jiné narostly do obří velikosti, jako například kytovec Livyatan melvillei, jenž měřil mezi 13,5 až 17,5 metry. Koncem miocénu, před asi 11 miliony lety, začalo významně klesat množství a rozmanitost vorvaňů, nicméně jiní draví kytovci, jako kosatky, během pliocénu postupně vyplňovali tuto uvolněnou ekologickou niku.[87][92]
Na základě fosilních důkazů se zdá, že jiné druhy žraloků, jako je například i žralok bílý, reagovaly na konkurenční tlak ze strany megalodonů tak, že začaly obývat převážně chladnější vody. V oblastech, kde společně koexistovaly, jako například během pliocénu u Baja California, možná střídavě žily během různých ročních období.[93][94] Podobně jako současné druhy žraloků, i megalodon se potom pravděpodobně uchyloval ke kanibalismu.[95]
Lovecké strategie

Žraloci často využívají k lovu velké kořisti různých způsobů. Megalodon mohl používat podobné lovecké techniky jako současný žralok bílý,[96] na základě otisků zubů ve velrybích fosiliích se však zdá, že způsobů, jak zabít kořist, využíval více než současný bílý žralok.[37]
Jedním konkrétním exemplářem je například devítimetrový pozůstatek miocenní kosticovité velryby neznámého taxonu, na němž lze analyzovat útočné chování megalodona. Na rozdíl od žraloků bílých, kteří se zaměřují na břicho kořisti, megalodon pravděpodobně útočil na srdce a plíce. Svými silnými zuby, pomocí kterých uměl překousnout i tvrdé kosti, prokousl kořisti hrudní koš a další odolné kosti. Způsob útoku se mohl také odlišovat na základě velikosti kořisti. Například fosilní pozůstatky menších kytovců, jako byli zástupci čeledi Cetotheriidae, naznačují, že do nich megalodon prudce narazil zespodu a tak jim způsobil kompresní zlomeninu.[37][96]
Během pliocénu se vyvinuly větší a vyspělejší druhy kytovců.[97] Megalodon si tedy asi vytvořil nové lovecké strategie, jak tyto větší velryby zabít. Byla nalezena řada zkamenělých kostí, které kdysi tvořily prsní ploutve či ocasní obratle, patřící pliocenním druhům velryb. Na těchto pozůstatcích byly objeveny otisky po zubech megalodonů, z čehož plyne, že tento žralok znehybnil větší velryby tím, že jim poškodil ploutve a teprve potom je zabil a sežral.[54][37]
Vývoj mláďat

Mláďata megalodona, stejně jako současných žraloků, žila ve „školkách“ s velkým množstvím potravy, které jim zároveň poskytovaly ochranu před predátory. Na základě fosilních důkazů se zdá, že se tyto areály nacházely v teplých vodách pobřežních oblastí, kde bylo jak méně nepřátel, tak dostatek jídla.[28] Tyto oblasti byly objeveny například v panamské formaci Gatun, v marylandské formaci Calvert, na Kanárských ostrovech či na Floridě (souvrství Bone Valley). Vzhledem k tomu, že všichni existující lamnovití (kam se megalodon někdy řadí) rodí živá mláďata, je možné, že se i u tohoto druhu vyvinula živorodost.[98][99] Podle jedněch odhadů mláďata měřila asi 3,5 metru,[53] další předpokládají, že mohla být jen asi dvoumetrová, což by je mohlo vystavovat hrozbě ze strany jiných druhů žraloků, jako byl kladivoun velký (Sphyrna mokarran) či vyhynulý miocenní druh Hemipristis serra.[28] Potrava mláďat se potom měnila s věkem.[100] Zprvu lovila ryby,[28] mořské želvy,[79]dugongy[101] či malé kytovce, na větší velryby se přeorientovala teprve v dospělosti.[90]
Nicméně ve fosilních pozůstatcích byly objeveny i případy, kdy mladí megalodoni občas napadli i mnohem větší kytovce než byli sami. Například na žebrech pliocenního keporkaka či plejtváka byly objeveny kousance od 4 až 7metrového žraloka, které vykazovaly známky hojení. Je možné, že velrybě toto zranění způsobil nedospělý megalodon.[102][103]
Vyhynutí
Klimatické změny
Během doby, v níž žil megalodon, Zemi postihla řada změn, které ovlivnily mořský život. Postupné ochlazování planety, které začalo v oligocénu před asi 35 miliony let, nakonec vedlo k zamrznutí pólů. Geologické události změnily pohyb mořských proudů a srážek, došlo k uzavření rovníkové cirkumglobální vodní cesty mezi Amerikami a napříč mořem Tethys, což vedlo k ochlazování oceánů. Zastavením Golfského proudu přestala zásobovat na živiny bohatá voda velké mořské ekosystémy, což mohlo způsobit úbytek megalodonovy potravy. Vzhledem k tomu, že se megalodon možná nevyskytoval v oblastech s chladnější vodou, pravděpodobně nebyl schopen udržet velké množství metabolického tepla, a proto se jeho areál výskytu mohl zmenšit na stále ubývající oblasti s teplou vodou[84][104][105] (tato teorie je nicméně sporná, viz níže).
Na základě fosilních důkazů se zdá, že v pliocénu, kdy výrazně poklesla teplota vody, megalodon vymizel z mnoha oblastí po celém světě.[93] Mezi pliocénem až pleistocénem před 5 miliony až 12 tisíci lety došlo k velkému kolísání mořské hladiny v důsledku rozšiřování ledovců na pólech, což mělo negativní dopad na pobřežní oblasti; tato událost mohla přispět k vyhynutí megalodona a několika jiných velkých mořských druhů.[106] Tyto změny mohly způsobit úbytek vhodných mělčin s teplou vodou, kde se vyvíjela mláďata;[104] oblastí klíčových pro přežití řady druhů žraloků, i proto že chrání jejich mláďata před útoky predátorů.[28][107]
Nicméně analýza výskytu megalodona v průběhu času ukazuje, že změna teploty nehrála přímou roli v jeho vyhynutí. Jeho rozšíření během miocénu a pliocénu nekoreluje s oteplováním a ochlazováním planety. I přestože populace megalodonů během pliocénu klesala, žralok byl ve skutečnosti schopen obývat i jiné než tropické oblasti. Jeho fosilie byly objeveny v místech, kde panovaly průměrné teploty mezi 12 až 27 °C, s celkovým rozpětím teplot 1 až 33 °C, což ukazuje, že rozsah vodních stanovišť nebyl výrazně ovlivněn změnami teploty, k nimž došlo. Výsledky rovněž potvrzují teorii, že tento žralok byl mezotermní.[14][56] Podle některých spekulací mohli tito obří žraloci i další velcí živočichové vyhynout asi před 2,6 miliony let vinou exploze supernovy ve vzdálenosti asi 150 světelných let od Země.[108] Ovšem novější datování vymření na dobu před 3,6 miliony lety tuto teorii vyvrací.[16]
Změny ekosystému
Mořští savci dosahovali největší diverzity během miocénu.[109] Například kosticovců tehdy existovalo více než dvacet rodů; v současnosti se jich vyskytuje pouze šest.[110] Tato rozmanitost kořisti byla pro superpredátory, mezi které se řadil i megalodon, ideální.[85] Na konci období miocénu však mnoho druhů kosticovců vyhynulo[87] a žijící druhy se mohly vyvinout v rychlejší plavce, což ztěžovalo jejich ulovení.[111] Navíc po uzavření průlivu ve Střední Americe poklesla rozmanitost a četnost tropických druhů velryb.[105] Období vyhynutí megalodona koreluje s poklesem počtu mnoha vývojových linií kosticovců, a je možné, že byl na nich, jako na zdroji potravy, závislý.[83]
Kromě toho během pliocénu vyhynulo 36 % velkých mořských druhů, včetně 55 % mořských savců, 35 % mořských ptáků, 9 % žraloků a 43 % mořských želv. Vyhynutí postihlo především živočichy endotermní a mezotermní, což ukazuje na nedostatek potravy (a zároveň podporuje teorii, že megalodon byl mezotermní).[56][106] Obřímu žralokovi nemusely stačit ztenčující se zdroje potravy.[104] Ochlazování během pliocénu mu pak mohlo zabránit v přístupu do polárních oblastí, kam se stěhovaly velké velryby.[71][105]
Rovněž konkurence ze strany nových predátorů, jako vorvaňů v miocénu, a kosatek a žraloků bílých v pliocénu,[87][92][112] mohla přispět k poklesu stavů megalodona. Nově vyvinutí kytovci živící se jinými velrybami se pak pravděpodobně mohli vyrovnat i se stále se rozšiřující studenou vodou a navíc obývali i tropické oblasti.[14][104][92][113]
Vyhynutí megalodona způsobilo další změny v mořských společenstvech.[114] Výrazně se zvětšili kosticovci, i když možná kvůli klimatickým změnám. Také je naopak možné, že zvětšení velikosti kosticovců způsobilo vyhynutí megalodona, protože ten zabíjel převážně menší druhy. Vyhynutí megalodona mělo také pozitivní dopad na další velké predátory, jako na velkého bílého žraloka, který obsadil oblasti uvolněné megalodonem.[14][83][112][115]
V kultuře a kryptozoologii

Megalodon je postavou některých děl včetně filmů a románů, v nichž je často zobrazován jako mořská příšera.[116][117] Tři jedinci (dva dospělci a mládě) megalodona byli představeni v dokumentu BBC Putování s dinosaury: Monstra pravěkých oceánů s Nigelem Marvenem, kde je popisován jako vrcholový predátor své doby.[118] V dokumentárním cyklu Jurské bojiště byl zobrazen, jak útočí na velryby Brygmophyseter.[119] Je rovněž součástí některých filmů, například Žralok útočí 3: Lidožrout, Megažralok vs. obří chobotnice, Megažralok versus crocosaurus[116] či DCU: Liga spravedlivých: Trůn Atlantidy.[120] V některých příbězích, jako Tedford and the Megalodon, je vykresleno znovuobjevení žraloka.[121] V románu Meg: Teror pochází z hlubin od Stevea Altena je žralok datován do období dinosaurů a na původním americkém přebalu je zobrazen, jak zabíjí tyranosaura; to je však nesmysl, neboť tato zvířata žila ve zcela jiných geologických obdobích.[122] Autor pak napsal ještě několik navazujících románů s megalodony.[116]
V roce 2013 natočil Discovery Channel televizní film Megalodon: Obří superžralok žije. Tento kontroverzní dokument předkládal údajné důkazy, které naznačovaly, že megalodon nevyhynul a stále existuje. Na film se snesla vlna kritiky vzhledem k tomu, že byl zcela smyšlený; například všechny údajné vědce představovali herci. O rok později tento pseudodokument znovu DC odvysílal společně s novými hodinovými filmy Megalodon: Nové důkazy a Žralok v hlubinách: Zuřivá ponorka, které opět vyvolaly kritiku médií i vědecké komunity.[37][123][124]
Z dvacátého století existuje rovněž několik údajných pozorování megalodonů, avšak neověřených. Podle těchto pozorování měli žraloci měřit mezi 10 a 90 metry. Je pravděpodobné, že šlo o chybně identifikované žraloky velrybí. Jeden polynéský mýtus hovoří o třicetimetrovém žralokovi, kterého nazývá pánem hlubin, pravděpodobně se však také jedná o žraloka velrybího. Zprávy o údajně nových zubech megalodona, které například objevila posádka lodi HMS Challenger roku 1873, a jež byly chybně datovány do období před 24 000 až 11 000 lety, jsou chybné a pravděpodobně se jedná o dobře zakonzervované zuby, které si při fosilizaci zachovaly bílou barvu (zuby megalodona mohou být zbarveny různými odstíny, od špinavě bílé přes tmavě hnědou až po šedou, a někdy dokonce vyzdvihnuty do mladší geologické vrstvy). Tvrzení, že by megalodon mohl dodnes přežívat v mořských hlubinách, podobně jako v sedmdesátých letech dvacátého století objevený žralok velkoústý (Megachasma pelagios), je nepravděpodobné, protože megalodon žil primárně v oblastech teplých pobřežních vod a pravděpodobně by nepřežil v chladném a na potravu chudém prostředí hlubokého moře.[71][125][126]
O tomto obřím žralokovi coby potenciálním kryptickém přežívajícím druhu psal v českém prostředí například popularizátor kryptozoologie Jaroslav Mareš (1937-2021).[127]
Odkazy
Poznámky
- ↑ Studie Henriho Cappety z roku 1974, zkoumající paleogenní žraloky, vytvořila podrod Megaselachus, ve kterém byl megalodon zařazen jako Otodus (Megaselachus) megalodon společně s druhem O. (M.) chubutensis. Revize z roku 2006 poté povýšila Megaselachus na rod, čímž by megalodon patřil do tohoto rodu jako Megaselachus megalodon.[29]
Reference
V tomto článku byl použit překlad textu z článku Megalodon na anglické Wikipedii.
- ↑ Kenshu Shimada, Harry M. Maisch IV, Victor J. Perez, Martin A. Becker & Michael L. Griffiths (2022). Revisiting body size trends and nursery areas of the Neogene megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), reveals Bergmann’s rule possibly enhanced its gigantism in cooler waters. Historical Biology. doi: https://doi.org/10.1080/08912963.2022.2032024
- ↑ Griffiths, M. L.; et al. (2023). Endothermic physiology of extinct megatooth sharks. Proceedings of the National Academy of Sciences. 120 (27): e2218153120. doi: 10.1073/pnas.2218153120
- ↑ KENDALL, Haven. 100 Greatest Science Discoveries of All Time. Westport: Libraries Unlimited, 1997. ISBN 978-1-59158-265-6. S. 25–26. (anglicky)
- ↑ HSU, Kuang-Tai. The Path to Steno's Synthesis on the Animal Origin of Glossopetrae. Boulder: Geological Society of America, 2009. Dostupné online. ISBN 978-0-8137-1203-1. Kapitola The Revolution in Geology from the Renaissance to the Enlightenment. (anglicky)
- ↑ EILPERIN, J. Demon Fish. [s.l.]: Pantheon Books, 2012. Dostupné online. ISBN 978-0-7156-4352-5. S. 43. (anglicky)
- ↑ AGASSIZ, L. Recherches sur les poissons fossiles. [s.l.]: [s.n.], 1843. Dostupné online. S. 41. (francouzsky)
- ↑ a b c d NYBERG, Kevin G.; CIAMPAGLIO, Charles N.; WRAY, Gregory A. Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth. Journal of Vertebrate Paleontology. 2006-12-11, roč. 26, čís. 4, s. 806–814. [806:TTAOTG2.0.CO;2 Dostupné online] [cit. 2017-12-29]. ISSN 0272-4634. DOI 10.1671/0272-4634(2006)26[806:ttaotg]2.0.co;2.
- ↑ KEYES, I. W. New records of the Elasmobranch C. Megalodon (Agassiz) and a review of the genus Carcharodon in the New Zealand fossil record. New Zealand Journal of Geology and Geophysics. 1972-06-01, roč. 15, čís. 2, s. 228–242. Dostupné online [cit. 2017-12-29]. ISSN 0028-8306. DOI 10.1080/00288306.1972.10421956.
- ↑ LIDDEL, Henry George. μέγας. [s.l.]: A Greek–English Lexicon
- ↑ LIDDEL, Henry George. ὀδoύς. [s.l.]: A Greek–English Lexicon
- ↑ a b AGUILERA, Orangel A.; GARCÍA, Luís; COZZUOL, Mario A. Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation. Paläontologische Zeitschrift. 2008-06-01, roč. 82, čís. 2, s. 204–208. Dostupné online [cit. 2017-12-29]. ISSN 0031-0220. DOI 10.1007/bf02988410. (anglicky)
- ↑ YABE, H; GOTO, M; KANEKO, N. Age of Carcharocles megalodon (Lamniformes: Otodontidae): A review of the stratigraphic records. The Palaeontological Society of Japan. 2004-03-01, s. 7–15. Dostupné online [cit. 2018-01-09].
- ↑ a b c d GOTTFRIED, Michael D.; FORDYCE, R. Ewan. An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships. Journal of Vertebrate Paleontology. 2001-12-14, roč. 21, čís. 4, s. 730–739. Dostupné online [cit. 2018-01-09]. ISSN 0272-4634. DOI 10.1671/0272-4634(2001)021[0730:aasoca]2.0.co;2.
- ↑ a b c d e f g h i PIMIENTO, Catalina; MACFADDEN, Bruce J.; CLEMENTS, Christopher F. Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms. Journal of Biogeography. 2016-08-01, roč. 43, čís. 8, s. 1645–1655. Dostupné online [cit. 2018-01-09]. ISSN 1365-2699. DOI 10.1111/jbi.12754. (anglicky)
- ↑ a b c d PIMIENTO, Catalina; CLEMENTS, Christopher F. When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record. PLOS ONE. 2014-10-22, roč. 9, čís. 10, s. e111086. Dostupné online [cit. 2018-01-09]. ISSN 1932-6203. DOI 10.1371/journal.pone.0111086.
- ↑ a b Giant 'megalodon' shark extinct earlier than previously thought. EurekAlert! [online]. [cit. 2019-02-13]. Dostupné online. (anglicky)
- ↑ a b c SHIMADA, Kenshu; CHANDLER, Richard E.; LAM, Otto Lok Tao. A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade. Historical Biology. 2017-07-04, roč. 29, čís. 5, s. 704–714. Dostupné online [cit. 2018-01-09]. ISSN 0891-2963. DOI 10.1080/08912963.2016.1236795.
- ↑ a b c d PIMIENTO, Catalina; BALK, Meghan A. Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators. Paleobiology. Roč. 41, čís. 03, s. 479–490. Dostupné online [cit. 2018-01-09]. DOI 10.1017/pab.2015.16.
- ↑ a b EHRET, Dana J.; HUBBELL, Gordon; MACFADDEN, Bruce J. Exceptional preservation of the white shark Carcharodon (Lamniformes, Lamnidae) from the early Pliocene of Peru. Journal of Vertebrate Paleontology. 2009-03-12, roč. 29, čís. 1, s. 1–13. Dostupné online [cit. 2018-01-09]. ISSN 0272-4634. DOI 10.1671/039.029.0113.
- ↑ KLIMLEY, Peter; AINLEY, David. Great white sharks : the biology of Carcharodon carcharias. San Diego: Academic Press, 1996. 517 s. ISBN 9780124150317. OCLC 392312939 S. 23–25. [Dále jen Klimley & Ainley].
- ↑ a b c RENZ, Mark. Megalodon : hunting the hunter. Lehigh Acres, Fla.: PaleoPress, 2002. 159 s. ISBN 9780971947702. OCLC 52125833 S. 30–31. [Dále jen Renz].
- ↑ Klimley & Ainley, s. 70.
- ↑ a b c d Megalodon Shark Evolution - Rersearch and Science - by Lutz Andres - Carcharodon versus Carcharocles. fossilguy.com. Dostupné online [cit. 2018-01-09]. (anglicky)
- ↑ Renz, s. 28–31.
- ↑ SIVERSON, Mikael. Late Cretaceous (Cenomanian-Campanian) mid-palaeolatitude sharks of Cretalamna appendiculata type. Acta Palaeontologica Polonica. Dostupné online [cit. 2018-01-09]. DOI 10.4202/app.2012.0137. (anglicky)
- ↑ Renz, s. 17.
- ↑ BENTON, M. J.; PEARSON, P. N. Speciation in the fossil record. Trends in Ecology & Evolution. 2001-07-01, roč. 16, čís. 7, s. 405–411. PMID: 11403874. Dostupné online [cit. 2018-01-09]. ISSN 1872-8383. PMID 11403874.
- ↑ a b c d e f g h PIMIENTO, Catalina; EHRET, Dana J.; MACFADDEN, Bruce J. Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama. PLOS ONE. 2010-05-10, roč. 5, čís. 5, s. e10552. Dostupné online [cit. 2018-01-09]. ISSN 1932-6203. DOI 10.1371/journal.pone.0010552.
- ↑ HENRI., Cappetta,. Chondrichthyes II : Mesozoic and Cenozoic Elasmobranchii. Stuttgart: G. Fischer Verlag 193 s. ISBN 9783899370461.
- ↑ a b Klimley & Ainley, s. 64–65.
- ↑ a b Renz, s. 35–36.
- ↑ Could Megalodon Have Looked Like a BIG Sandtiger?. www.elasmo-research.org [online]. [cit. 2018-01-10]. Dostupné online.
- ↑ The Megalodon was more slender than depicted in movies, study shows. phys.org [online]. [cit. 2024-01-22]. Dostupné online.
- ↑ Renz, s. 87.
- ↑ PORTELL, Roger W.; HUBBELL, Gordon; DONOVAN, Stephen K. Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles. Caribbean Journal of Science. 2008-01-01, roč. 44, čís. 3, s. 279–286. Dostupné online [cit. 2018-01-10]. ISSN 0008-6452. DOI 10.18475/cjos.v44i3.a2.
- ↑ a b Klimley & Ainley, s. 57.
- ↑ a b c d e f g h PROTHERO, Donald. The story of life in 25 fossils : tales of intrepid fossil hunters and the wonders of evolution. New York: [s.n.], 2015. 389 s. ISBN 9780231171908. OCLC 900031557 S. 96–110.
- ↑ HELFMAN, Gene; BURGESS, George. Sharks : the animal answer guide. Baltimore: Hopkins University Press, 2014. Dostupné online. ISBN 9781421413105. OCLC 877868297 S. 19.
- ↑ RANDALL, John E. Size of the Great White Shark (Carcharodon). Science. 1973-07-13, roč. 181, čís. 4095, s. 169–170. PMID: 17746627. Dostupné online [cit. 2018-01-10]. ISSN 0036-8075. DOI 10.1126/science.181.4095.169. PMID 17746627. (anglicky)
- ↑ Klimley & Ainley, s. 99.
- ↑ SCHEMBRI, Patrick. MALTA'S NATURAL HERITAGE. Natural Heritage. 1994-01-01. Dostupné online [cit. 2018-01-10].
- ↑ PAPSON, Stephen. “Cross the Fin Line of Terror”: Shark Week on the Discovery Channel. Journal of American Culture. 1992-12-01, roč. 15, čís. 4, s. 67–81. Dostupné online [cit. 2018-01-10]. ISSN 1542-734X. DOI 10.1111/j.1542-734x.1992.1504_67.x. (anglicky)
- ↑ Klimley & Ainley, s. 60.
- ↑ Klimley & Ainley, s. 55–60.
- ↑ Renz, s. 88.
- ↑ SHIMADA, Kenshu. The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae). Journal of Fossil Research. 2002-01-01, roč. 35, s. 28–33. Dostupné online [cit. 2018-01-10].
- ↑ Kenshu Shimada (2019). The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited. Historical Biology (advance online publication). doi: 10.1080/08912963.2019.1666840
- ↑ https://resources.depaul.edu/newsroom/news/press-releases/Pages/megatooth-shark.aspx
- ↑ Cooper, Jack A.; et al. (2020). Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction. Scientific Reports. doi: 10.1038/s41598-020-71387-y
- ↑ True size of prehistoric mega-shark finally revealed. phys.org [online]. [cit. 2020-09-15]. Dostupné online. (anglicky)
- ↑ Victor J. Perez, Ronny M. Leder, and Teddy Badaut (2021). Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions. Palaeontologia Electronica. 24 (1): a09. doi: https://doi.org/10.26879/1140
- ↑ Kenshu Shimada, Martin A. Becker & Michael L. Griffiths (2020). Body, jaw, and dentition lengths of macrophagous lamniform sharks, and body size evolution in Lamniformes with special reference to 'off-the-scale' gigantism of the megatooth shark, Otodus megalodon. Historical Biology. doi: https://doi.org/10.1080/08912963.2020.1812598
- ↑ a b c Klimley & Ainley, s. 61.
- ↑ a b c d WROE, S. & kol. Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite. Journal of Zoology. 2008, s. 336–342. DOI 10.1111/j.1469-7998.2008.00494.x.
- ↑ JACOBY, David M. P.; SIRIWAT, Penthai; FREEMAN, Robin. Is the scaling of swim speed in sharks driven by metabolism?. Biology Letters. 2015-12-01, roč. 11, čís. 12, s. 20150781. PMID: 26631246. Dostupné online [cit. 2018-01-10]. ISSN 1744-9561. DOI 10.1098/rsbl.2015.0781. PMID 26631246. (anglicky)
- ↑ a b c FERRÓN, Humberto G. Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks. PLOS ONE. 2017-09-22, roč. 12, čís. 9, s. e0185185. Dostupné online [cit. 2018-01-10]. ISSN 1932-6203. DOI 10.1371/journal.pone.0185185.
- ↑ a b Klimley & Ainley, s. 55.
- ↑ a b BENDIX-ALMGREEN, S. E. Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn. Bulletin of the Geological Society of Denmark. 1983, s. 1–32.
- ↑ REOLID, M.; MOLINA, J. M. Registro de Carcharocles megalodon en el sector oriental de la Cuenca del Guadalquivir (Mioceno superior, Sur de España). Estudios Geológicos. 2015-12-30, roč. 71, čís. 2, s. 032. Dostupné online [cit. 2018-01-10]. ISSN 1988-3250. DOI 10.3989/egeol.41828.342. (anglicky)
- ↑ a b c Renz, s. 33.
- ↑ CRANE, Brent. A Prehistoric Killer, Buried in Muck. The New Yorker. 2017-03-05. Dostupné online [cit. 2018-01-10]. ISSN 0028-792X.
- ↑ For Sale: World's Largest Shark Jaws. Live Science. Dostupné online [cit. 2018-01-10].
- ↑ UYENO, T. & kol. The Description of an Almost Complete Tooth Set of Carcharodon megalodon from a Middle Miocene Bed in the Saitama Prefecture, Japan. Saitama Museum of Natural History Bulletin. 1989, s. 73–85.
- ↑ Klimley & Ainley, 20–21.
- ↑ a b Renz, s. iv.
- ↑ Renz, s. 1, 129.
- ↑ Klimley & Ainley, s. 63.
- ↑ ANDERSON, Philip S. L.; WESTNEAT, Mark W. Feeding mechanics and bite force modelling of the skull of Dunkleosteus terrelli, an ancient apex predator. Biology Letters. 2007-02-22, roč. 3, čís. 1, s. 77–80. Dostupné online [cit. 2018-01-10]. ISSN 1744-9561. DOI 10.1098/rsbl.2006.0569. (anglicky)
- ↑ ANDERSON, Philip S. L.; WESTNEAT, Mark W. A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi). Paleobiology. Roč. 35, čís. 02, s. 251–269. Dostupné online [cit. 2018-01-10]. DOI 10.1666/08011.1.
- ↑ Megalodon Shark Facts and Information. www.fossilguy.com. Dostupné online [cit. 2018-01-10]. (anglicky)
- ↑ a b c d ROESCH, B. S. A Critical Evaluation of the Supposed Contemporary Existence of Carcharocles megalodon. The Cryptozoology Review [online]. 1988 [cit. 2018-01-10]. Dostupné v archivu pořízeném z originálu.
- ↑ Klimley & Ainley, s. 63–65.
- ↑ STRINGER, G. L.; KING, L. Late Eocene Shark Coprolites from the Yazoo Clay in Northeastern Louisiana. New Mexico Museum of Natural History and Science, Bulletin 57. Vertebrate Corpolites: 301.. 2012.
- ↑ Klimley & Ainley, s. 56.
- ↑ PIMIENTO, Catalina; GONZÁLEZ-BARBA, Gerardo; EHRET, Dana J. Sharks and rays (Chondrichthyes, Elasmobranchii) from the late Miocene Gatun Formation of Panama. Journal of Paleontology. 2013/09, roč. 87, čís. 5, s. 755–774. Dostupné online [cit. 2018-01-11]. ISSN 0022-3360. DOI 10.1666/12-117.
- ↑ Klimley & Ainley, s. 67.
- ↑ FITZGERALD, Erich. A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia. Memoirs of the Museum of Victoria. 2004-01-01, roč. 61. Dostupné online [cit. 2018-01-11]. DOI 10.24199/j.mmv.2004.61.12.
- ↑ a b Klimley & Ainley, s. 78.
- ↑ a b c AGUILERA, Orangel; RODRIGUES, Dione; R. DE AGUILERA, D. Giant-toothed white sharks and wide-toothed mako (Lamnidae) from the Venezuela Neogene: Their role in the Caribbean, shallow-water fish assemblage. Caribbean Journal of Science. 2004-12-01, roč. 40. Dostupné online [cit. 2018-01-11].
- ↑ Fossilworks: Carcharocles megalodon. fossilworks.org [online]. [cit. 2018-01-11]. Dostupné v archivu pořízeném dne 2018-01-11.
- ↑ Klimley & Ainley, s. 71–75.
- ↑ MARTIN, Jeremy E.; TACAIL, Théo; ADNET, Sylvain. Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs. Chemical Geology. Roč. 415, s. 118–125. Dostupné online [cit. 2018-01-11]. DOI 10.1016/j.chemgeo.2015.09.011.
- ↑ a b c d COLLARETA, Alberto; LAMBERT, Olivier; LANDINI, Walter. Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru. Palaeogeography, Palaeoclimatology, Palaeoecology. Roč. 469, s. 84–91. Dostupné online [cit. 2018-01-11]. DOI 10.1016/j.palaeo.2017.01.001.
- ↑ a b MORGAN, Gary S. Whither the giant white shark?. Paleontology Topics. 1994, s. 1–2.
- ↑ a b Klimley & Ainley, s. 75.
- ↑ LANDINI, Walter; ALTAMIRANO-SIERRA, Alì; COLLARETA, Alberto. The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru). Journal of South American Earth Sciences. Roč. 73, s. 168–190. Dostupné online [cit. 2018-01-11]. DOI 10.1016/j.jsames.2016.12.010.
- ↑ a b c d e f g LAMBERT, Olivier; BIANUCCI, Giovanni; POST, Klaas. The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru. Nature. 2010/07, roč. 466, čís. 7302, s. 105–108. Dostupné online [cit. 2018-01-11]. ISSN 1476-4687. DOI 10.1038/nature09067. (En)
- ↑ COMPAGNO, Leonard J. V. Alternative life-history styles of cartilaginous fishes in time and space. Environmental Biology of Fishes. 1990-08-01, roč. 28, čís. 1–4, s. 33–75. Dostupné online [cit. 2018-01-11]. ISSN 0378-1909. DOI 10.1007/bf00751027. (anglicky)
- ↑ FERRETTI, Francesco; WORM, Boris; BRITTEN, Gregory L. Patterns and ecosystem consequences of shark declines in the ocean. Ecology Letters. 2010-08-01, roč. 13, čís. 8, s. 1055–1071. Dostupné online [cit. 2018-01-11]. ISSN 1461-0248. DOI 10.1111/j.1461-0248.2010.01489.x. (anglicky)
- ↑ a b Klimley & Ainley, s. 74–75.
- ↑ BIANUCCI, Giovanni; LANDINI, Walter. Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy. Zoological Journal of the Linnean Society. 2006-09-01, roč. 148, čís. 1, s. 103–131. Dostupné online [cit. 2018-01-11]. ISSN 0024-4082. DOI 10.1111/j.1096-3642.2006.00228.x.
- ↑ a b c HEYNING, John E.; DAHLHEIM, Marilyn E. Orcinus orca. Mammalian Species. 1988-01-15, čís. 304, s. 1–9. Dostupné online [cit. 2018-01-11]. ISSN 0076-3519. DOI 10.2307/3504225.
- ↑ a b Klimley & Ainley, s. 77.
- ↑ Paleoecology of Megalodon and the White Shark. www.elasmo-research.org [online]. [cit. 2018-01-11]. Dostupné online.
- ↑ TANKE, Darren; J CURRIE, Philip. Head-biting behavior in theropod dinosaurs: Paleopathological evidence. Gaia. 2000-01-01, roč. 15. Dostupné online [cit. 2018-01-11].
- ↑ a b GODFREY, S. J.; ALTMAN, J. A Miocene Cetacean Vertebra Showing a Partially Healed Compression Factor, the Result of Convulsions or Failed Predation by the Giant White Shark, Carcharodon megalodon. Jeffersoniana. 2005, s. 1–12.
- ↑ DEMÉRÉ, Thomas A.; BERTA, Annalisa; MCGOWEN, Michael R. The Taxonomic and Evolutionary History of Fossil and Modern Balaenopteroid Mysticetes. Journal of Mammalian Evolution. 2005-06-01, roč. 12, čís. 1–2, s. 99–143. Dostupné online [cit. 2018-01-11]. ISSN 1064-7554. DOI 10.1007/s10914-005-6944-3. (anglicky)
- ↑ PRESS, Europa. Identifican en Canarias fósiles de 'megalodón', el tiburón más grande que ha existido. europapress.es. 2013-09-02. Dostupné online [cit. 2018-01-12]. (španělsky)
- ↑ DULVY, Nicholas K.; REYNOLDS, John D. Evolutionary transitions among egg–laying, live–bearing and maternal inputs in sharks and rays. Proceedings of the Royal Society of London B: Biological Sciences. 1997-09-22, roč. 264, čís. 1386, s. 1309–1315. Dostupné online [cit. 2018-01-12]. ISSN 0962-8452. DOI 10.1098/rspb.1997.0181. (anglicky)
- ↑ Klimley & Ainley, s. 65.
- ↑ Renz, s. 129.
- ↑ GODFREY, S. The Ecphora. The Newsletter of Calvert Marine Museum Fossil Club. 2004, s. 1–13. Dostupné v archivu pořízeném z originálu. Archivováno 10. 12. 2010 na Wayback Machine.
- ↑ KALLAL, R. J.; GODFREY, S. J.; ORTNER, D. J. Bone reactions on a pliocene cetacean rib indicate short-term survival of predation event. International Journal of Osteoarchaeology. 2012-05-01, roč. 22, čís. 3, s. 253–260. Dostupné online [cit. 2018-01-12]. ISSN 1099-1212. DOI 10.1002/oa.1199. (anglicky)
- ↑ a b c d Extinction of Megalodon. www.elasmo-research.org [online]. [cit. 2018-01-12]. Dostupné online.
- ↑ a b c ALLMON, Warren D.; EMSLIE, Steven D.; JONES, Douglas S. Late Neogene Oceanographic Change along Florida's West Coast: Evidence and Mechanisms. The Journal of Geology. 1996-03-01, roč. 104, čís. 2, s. 143–162. Dostupné online [cit. 2018-01-12]. ISSN 0022-1376. DOI 10.1086/629811.
- ↑ a b PIMIENTO, Catalina; GRIFFIN, John N.; CLEMENTS, Christopher F. The Pliocene marine megafauna extinction and its impact on functional diversity. Nature Ecology & Evolution. 2017/08, roč. 1, čís. 8, s. 1100–1106. Dostupné online [cit. 2018-01-12]. ISSN 2397-334X. DOI 10.1038/s41559-017-0223-6. (En)
- ↑ Prehistoric Shark Nursery Spawned Giants. news.discovery.com [online]. 2012-03-10 [cit. 2018-01-12]. Dostupné v archivu pořízeném z originálu.
- ↑ http://www.osel.cz/10258-vyhubila-megalodony-supernova-na-konci-pliocenu.html
- ↑ Klimley & Ainley, s. 71.
- ↑ DOOLEY, Alton C. Jr; FRASER, Nicholas C.; LUO, Zhe-Xi. The earliest known member of the rorqual—gray whale clade (Mammalia, Cetacea). Journal of Vertebrate Paleontology. 2004-06-11, roč. 24, čís. 2, s. 453–463. Dostupné online [cit. 2018-01-12]. ISSN 0272-4634. DOI 10.1671/2401.
- ↑ Renz, s. 46.
- ↑ a b TELLES ANTUNES, Miguel; BALBINO, Ausenda. The great white shark Carcharodon carcharias (linné, 1758) in the Pliocene of Portugal and its early distribution in eastern Atlantic. Revista Espanola de Paleontologia. 2010-01-01, roč. 25, s. 1–6. Dostupné online [cit. 2018-01-12].
- ↑ Renz, s. 46–47.
- ↑ SLATER, Graham J.; GOLDBOGEN, Jeremy A.; PYENSON, Nicholas D. Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics. Proc. R. Soc. B. 2017-05-31, roč. 284, čís. 1855, s. 20170546. PMID: 28539520. Dostupné online [cit. 2018-01-12]. ISSN 0962-8452. DOI 10.1098/rspb.2017.0546. PMID 28539520. (anglicky)
- ↑ ADNET, Sylvain; BALBINO, Ausenda C.; ANTUNES, Miguel Telles. New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean. Neues Jahrbuch f??r Geologie und Pal??ontologie - Abhandlungen. 2010-04-01, roč. 256, čís. 1, s. 7–16. Dostupné online [cit. 2018-01-12]. DOI 10.1127/0077-7749/2009/0029.
- ↑ a b c WEINSTOCK, A. J. The Ashgate encyclopedia of literary and cinematic monsters. Farnham: Routledge, 2014. 640 s. Dostupné online. ISBN 9781409425625. OCLC 864899715 S. 107–108.
- ↑ RICK., Emmer,. Megalodon : fact or fiction?. New York: Infobase Publishing, 2010. 103 s. ISBN 9781438132105. OCLC 649838860 S. 80–85.
- ↑ Sea Monsters: A Walking with Dinosaurs Trilogy [online]. BBC, 2003. Kapitola 3.
- ↑ Jurassic Fight Club [online]. History Channel, 2008. Kapitola 5.
- ↑ SPAULDING, Ethan. Justice League: Throne of Atlantis [online]. Warner Bros. Animation.
- ↑ SHEPARD, J. McSweeney's mammoth treasury of thrilling tales. New York: Knopf Doubleday Publishing Group, 2007. 479 s. ISBN 9780307426826. OCLC 636328662 S. 9.
- ↑ ALTEN, S. Meg: A Novel of Deep Terror. Portland: Gere Donovan Press, 2011. ISBN 978-1-936666-21-8.
- ↑ BRIAN,, Winston,. The act of documenting : documentary film in the 21st century. New York, NY: [s.n.] 271 s. ISBN 9781501309182. OCLC 959372509
- ↑ FLANAGIN, Jake. Sorry, Fans. Discovery Has Jumped the Shark Week.. Op-Talk. Dostupné online [cit. 2018-01-12]. (anglicky)
- ↑ RICK., Emmer,. Megalodon : fact or fiction?. New York: Chelsea House 103 s. ISBN 9781438132105. OCLC 649838860 S. 80–85.
- ↑ Does Megalodon Still Live?. www.elasmo-research.org [online]. [cit. 2018-01-12]. Dostupné online.
- ↑ https://dinosaurusblog.com/2021/05/07/vzpominka-na-jaroslava-marese/
Literatura
- NOVÁK, Jan A. Megalodon. Pátrání po nejstrašnějším mořském predátorovi. [s.l.]: XYZ, 2011. 308 s. ISBN 978-80-7388-535-9.
- Kenshu Shimada, Matthew F. Bonnan, Martin A. Becker & Michael L. Griffiths (2021). Ontogenetic growth pattern of the extinct megatooth shark Otodus megalodon--implications for its reproductive biology, development, and life expectancy. Historical Biology. doi: https://doi.org/10.1080/08912963.2020.1861608
- Jack A. Cooper, John R. Hutchinson, David C. Bernvigeremy Cliff, Rory P. Wilson, Matt L. Dicken, Jan Menzel, Stephen Wroe, Jeanette Pirlo & Catalina Pimiento (2022). The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling. Science Advances. 8 (33): eabm9424. doi: 10.1126/sciadv.abm9424
Externí odkazy
- Obrázky, zvuky či videa k tématu Megalodon na Wikimedia Commons
- Taxon Carcharocles megalodon ve Wikidruzích
- Článek o vědeckém výzkumu druhu C. megalodon (anglicky)
- Článek o megalodonovi na webu Prehistoric Beast of the Week (anglicky)
- Článek o megalodonovi na webu časopisu 100+1 (česky)
